Gehirntheorie des Menschen
ISBN 978-3-00-068559-0
Monografie von Dr. rer. nat. Andreas Heinrich Malczan
10. Das Modul des Rotationsspeichers im limbischen System
Bereits in meiner Monografie „Das limbische System“ ISBN 978-3-00-037458-6 (deutsch) bzw. ISBN 978-3-00-042153-2 (englisch) habe ich das limbische System erklärt und eine Hypothese über seine Entstehung entworfen. Das oberste Segment des uralten Strickleitersystems war das olfaktorische. Die Signale der Geruchsrezeptoren trafen im sensorischen Zentrum des ersten Segments ein. Über Neuronen der Klasse 3 zogen sie zum motorischen Zentrum und aktivierten dort die Neuronen der Klasse 5, die ihrerseits die Motoneuronen des Körpers aktivierten. So wurden olfaktorische Signale zur Körpersteuerung eingesetzt, um Futter zu finden. Doch die Duftsignale verflüchtigten sich im Wasser – wo alle Tiere damals lebten – recht schnell. Dann war das olfaktorische Signal weg. Günstig war es, die Dauer des olfaktorischen Signals zu verlängern. Hier fand die Natur folgende Lösung.
.
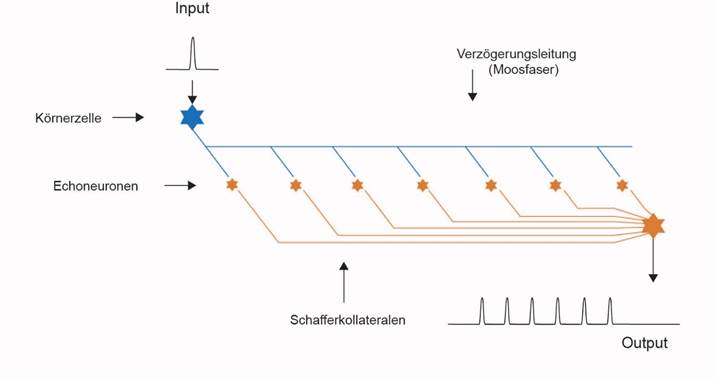
Abbildung 49: Prinzipieller Aufbau des Hippocampus am Beispiel einer MoosfaserDie Neuronen der
Klasse 3 bildeten jeweils ein Axon, die sogenannte Moosfaser, welches
in
regelmäßigen Abständen Axonkollateralen
bildete, die ihrerseits neue Neuronen
der Klasse 3 generierten. Diese Neuronen waren Echoneuronen, die vom
vorbeiziehenden Aktionspotential auf der Moosfaser aktiviert wurden und
so
selbst ein Aktionspotential bildeten. Da die Moosfasern keine
Myelinhülle
besaßen, breiteten sie die Aktionspotentiale relativ langsam
aus. Die Axone der
Echoneuronen – die Schaffer-Kollateralen – endeten
pro Moosfaser auf einem
eigenen Outputneuron. So wurde jedes Aktionspotential eines
olfaktorischen
Rezeptors in eine Folge von Aktionspotentialen umgewandelt, die nun
deutlich
länger dauerte. Die Länge der Moosfasern nahm zu, da
dies vorteilhaft war. So
konnte das Tier länger auf olfaktorische Reize reagieren, auch
wenn diese nur
kurz andauerten.
Die entstandene
Struktur war der Hippocampus. Sein Output erreichte auf der motorischen
Seite
den dortigen motorischen Kern, die Amygdala. Als diese zum Hippocampus
zurückprojizierte, entstand eine geschlossene Signalschleife.
In dieser konnte ein
einmal erzeugtes olfaktorisches Signal praktische unendlich lange
rotieren. So
wurde eine längere Beutesuche möglich. War die Beute
gefressen worden, konnte
ein Hemmungssignal aus den Verdauungsorganen in der Amygdala hemmende
Interneuronen aktiveren, die die ewige Rotation des Geruchssignals
stoppten.
Auch andere
Substrukturen des Nervensystems konnten in der Amygdala die hemmenden
Neuronen
ansteuern und die Signalrotation beenden. Andererseits konnten ebenso
erregende
Signale eine neue Signalrotation anstoßen, so dass nun ein
Rotationsgedächtnis
für Signale aller Art zur Verfügung stand.
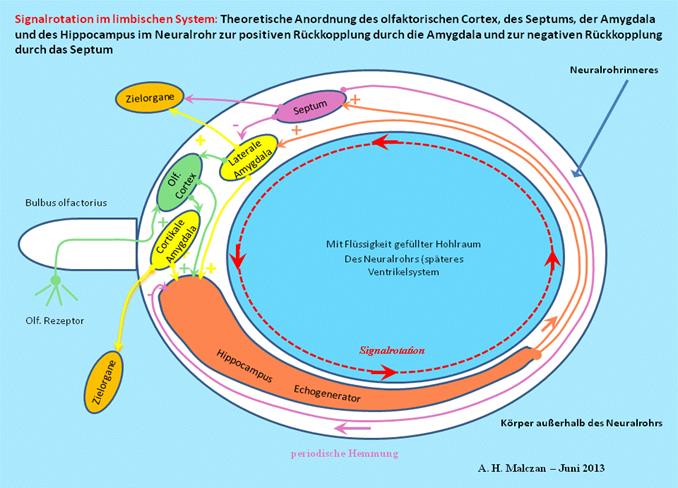
Abbildung 50: Prinzip der Signalrotation im limbischen System nach A. Malczan, 2013
Das Septum stellte in der ersten Etage des Strickleitersystems den Mittelwertkern dar. Seine Neuronen verwendeten bereits damals Acetylcholin und aktivierten damit die Muskeln. Noch heute dient dieser Neurotransmitter der Aktivierung der Muskeln. Damit die ununterbrochene Signalrotation nicht zur Überlastung der beteiligten Neuronen führte, entwickelte sich eine hemmende Rückprojektion vom Septum zur Amygdala, so dass die ständige Signalrotation für die Zeit unterbrochen wurde, die ein Signal brauchte, um von der Amygdala über den Hippocampus zum Septum zu gelangen. War dieses letzte Aktionspotential aufgebraucht, begann die Signalrotation erneut, denn in der geschlossenen Schleife gab es mehr Aktionspotentiale, als das Septum hemmen konnte. Der Hippocampus wurde so zum Rotationsspeicher des Nervensystems. Wenn ein Signal in den Rotationsspeicher Eingang gefunden hatte, stand es solange zur Verfügung, bis es durch einen weiteren Algorithmus gelöscht wurde.
Beispielsweise wurden
alle Retinasignale in den Hippocampus eingespeist. Dort rotierte dann
ein
komplettes Retinaabbild in Schleifen, für jedes Bildpixel und
jede visuelle
Modalität (Helligkeit, Farbe) gab es genau eine geschlossene
Signalschleife.
Die im Hippocampus
zwischengespeicherten Signale erreichten jedoch nicht direkt das
Bewusstsein,
welches die aktuelle Gegenwart auswertete. Die Vergangenheit wurde dem
Unterbewusstsein zur Verfügung gestellt. Dieses befand sich im
Cerebellum.
Daher erreichte der
limbische Rotationsoutput bei jedem Rotationsdurchlauf auch
über den anterioren
Thalamus das Cerebellum. Dort stehen die Rotationssignale
ständig zur
Verfügung. Und über das Cerebellum kehrten sie auch
zu ihren Ursprungsorten im
Cortex zurück. Sie waren ja – damals in
früher Evolutionszeit – mit den Originalsignalen
identisch.
Viele Jahrmillionen
später tauchte jedoch ein besonderes Phänomen auf.
Wenn man die Augen
schloss (oder alternativ ein weißes Blatt Papier anschaute),
so erschienen die
Rotationssignale des limbischen Systems vor dem geistigen Auge
– diesmal jedoch
in der Komplementärfarbe. Warum ist dies so?
Dies ist eine Folge
der Signaldivergenz im Nucleus olivaris, die im Lauf der Evolution
entstand.
Sie führte wahrscheinlich zu Veränderungen in den
Signalen.
Der Output der Retina
wird direkt zum limbischen System gesandt und nimmt dort an der
Signalrotation
teil, bei jedem Umlauf erreicht er über den anterioren
Thalamus auch das
Cerebellum über den Nucleus olivaris.
Als sich im Nucleus
olivaris auch eine Signaldivergenz etablierte, wurden die Signale im
Divergenzmodul der Olive genauso divergent behandelt wir im cortikalen
Divergenzmodul. Die Farbe beispielsweise wurde als maximumcodierter
Neuronenvektor verschlüsselt.
Allerdings entstand
ein Problem: Der Nucleus dentatus invertierte den Cerebellumoutput.
Dadurch
wurde er zu einem minimumcodierten Signalvektor. Diese Signalinversion
überführte die aktuelle Farbe in die
Komplementärfarbe. Diese erreicht – wenn
die Augen geschlossen werden oder ein weißes Blatt Papier
gesehen wird – wieder
über den Thalamus den Cortex und ruft dort die wahrgenommene
Komplementärfarbe
hervor. Grund ist die Signalinversion im Nucleus dentatus und die
Signaldivergenz im Nucleus olivaris, die es in früher Urzeit
noch nicht gab.
Diese Falschfarbenbilder sind ein Tribut an die Weiterentwicklung des
Spinocerebellums zur Inversionsschaltung.
Nachbilder in
Komplementärfarbe sind ein Indiz für die Richtigkeit
meiner Theorie.
Die Rotationssignale
des limbischen Systems erreichen vorwiegend das Cerebellum, welches im
Gehirnsystem das Unterbewusstsein repräsentiert. Dort
können die limbischen
Signale, die meist die Vergangenheit repräsentieren, die auch
deutlich länger
zurückliegen kann, mit Gegenwartssignalen kombiniert werden
und so neue
Komplexsignale bilden. Dies ist die Grundlage für unser
Langzeitgedächtnis.
Mehr kann hier nicht
dargestellt werden, weil die vier Wochen Zeit, in der meine Frau zur
Kur war,
nun fast zu Ende sind. Es bleibt nur noch Zeit für das
Korrekturlesen.
Das letzte Kapitel
zum Spracherwerb und zur Sprachsteuerung wird jedoch noch
fertiggestellt,
jedoch nur in Kurzform.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan