Gehirntheorie des Menschen
ISBN 978-3-00-068559-0
Monografie von Dr. rer. nat. Andreas Heinrich Malczan
11. Die Weiterentwicklung des Leiter-Nervensystem beim Menschen
In dieser Monografie wurde oft gezeigt, dass die Signalverarbeitung des Gehirns durch Module realisiert wird. Diese erzeugen als Output häufig Submodalitäten, die durch die Aufspaltung von ursprünglichen Modalitäten entstanden sind. So beispielsweise ging aus den Modalitäten Hell und Dunkel die neue Submodalität der Grauwerte hervor. Aus den Modalitäten Grün und Rot ging die Submodalität der Mischfarben Rot/Grün hervor, ebenso entstand eine Submodalität für die Mischfarben Rot/Blau und Blau/Grün. Diese drei Submodalitäten können wir zur neuen Submodalität Farbe zusammenfassen, da die drei Module im Gehirn pro Bildpixel genau übereinander liegen, darunter das Helligkeitsmodul.
Im Divergenzmodul mit räumlicher Signalausbreitung entstand die neue Modalität der orientierten Linienelemente, sowohl für die Submodalität der Farbe als auch für die Submodalität der Grauwerte. Aus ihr gingen im Zusammenwirken mit den Basalganglien die Modalität der bewegten Linienelemente sowohl für Farben als auch für Grautöne hervor.
Im Gehirn des Menschen, aber wahrscheinlich schon viel früher im Nervensystem der Wirbeltiere, entwickelte sich eine Tendenz, verschiedene Modalitäten zu trennen. Dies wurde im Verlauf der Evolution auch auf die Submodalitäten übertragen. Sie stellten ja neue, vorher nicht gekannte Signale dar.
Natürlich entstanden diese neuen Signale nicht durch Rezeptoren, jedoch waren sie dennoch der Output von Neuronen in den verschiedenen Modulen des Strickleiter-Nervensystems bzw. des Nachfolgersystems.
Wir erinnern uns: Die meisten Module – etwa im visuellen Bereich – entstanden im Prozess der Signaldivergenz. Output wurde auf mehrere, später auf viele Neuronen verteilt. Dies diente der Erhöhung der Ausfallsicherheit, stellte eine Art Bildung von Ersatzneuronen dar. Es ist unsinnig, anzunehmen, dies wäre erst im Cortex höherer Wirbeltiere passiert. Nein, dieser Prozess kann in der Evolution bereits sehr früh begonnen haben. Langsam, geradezu schleichend wurde einfach die Anzahl der Outputneuronen in den sensorischen und motorischen Zentren erhöht. Zusammen mit der Erhöhung der Rezeptorenarten und der Rezeptorenanzahl führte das in den Kopfsegmenten zu einer räumlichen Ausdehnung. Das dortige Kopfskelett – sobald vorhanden – erwarb wahrscheinlich die Fähigkeit, sich bei zunehmender Neuronenanzahl in seinem Inneren einfach auszudehnen. Noch heute ermöglichen die Fontanellen bei Neugeborenen die Ausdehnung des Gehirns in der ersten zwei Lebensjahren. Wir gehen hier davon aus, dass dieses Kopfskelett bereits vorhanden war.
Als die Schichtdicke der sensorischen und motorischen Zentren langsam wuchs, wodurch auch automatisch infolge der Signaldivergenz die Divergenzmodule entstanden, gab es neuen Output von den dadurch entstandenen Submodalitäten (Helligkeit/Farbe/Orientierung von Linien usw.).
Bei denjenigen segmentierten Bilateria, deren Linien zu den Primaten und zum Homo sapiens führten, begann ein völlig neues, bisher nicht dagewesenes Konzept zu greifen:
Der Output der neuen Submodalitäten aus den verschiedenen Modulen der ersten Etage wurde dem Rezeptoroutput gleichgestellt.
Er wurde so behandelt, als sei eine neue Rezeptorenart entstanden, die diesen Output im obersten Leitersegment ablieferte.
Dies hatte weitreichende Folgen. Im Leitersystem der Kopfsegmente beanspruchte jede Modalität ein eigenes Leitersegment. Als die neuen Submodalitäten sich entwickelten, mussten sie deshalb ebenso ein eigenes Leitersegment beanspruchen. So entstand über dem bisherigen obersten, cortikalen Leitersegment ein neues Leitersegment für die Signale der neuen Submodalitäten. Quasi ein Cortex über dem Cortex. Die Signale der neuen Submodalitäten wurden nach oben weitergegeben.
Technisch war dies jedoch schwierig umzusetzen. In der Höhe konnte kein neues Leitersegment entstehen, der verfügbare Raum wurde von den vorhandenen Neuronen und Axonen komplett ausgefüllt und war nach oben bereits durch den Schädelknochen begrenzt (auch wenn der vielleicht damals noch aus Knorpel bestanden haben mag).
So blieb den neuen Leitersegmenten nur das Ausweichen zur Seite. Das nächste Leitersegment bildete sich daher seitlich neben dem cortikalen Leitersegment. Zwischen dem bisherigen cortikalen Segment und dem neuen Segment gab es jedoch die klassischen Verbindungen.
Vielleicht wurde diese Entwicklung durch eine Eigenart der Neuronenklasse 4 bewirkt. Sie spezialisierten sich im Verlauf der Evolution darauf, neue Signale zum nächsten Nachbarsegment weiterzuleiten. Gab es kein übergeordnetes Segment, so schufen sie selbst ein neues, indem ihre Axone dem vorhandenen Gradientengefälle eines vorhandenen Kopfmarkers folgten und vorhandene Proneuronen der Klasse 3 aktivierten, so dass diese zu Projektionsneuronen der Klasse 3 wurden. Deren Axone zogen – gesteuert vom motorischen Marker der betreffenden Körperseite – zur motorischen Seite und generierten neue Projektionsneuronen der Klasse 5, die wiederum abwärts projizierten. Damit war eine neue Etage, also ein neues Segment im Nervensystem entstanden.
Es war der gleiche Vorgang wie bei der Entstehung von neuen Rezeptoren. Deren Signale generierten im jeweiligen Segment letztlich auch die Neuronen der Klasse 4, die sie an die übergeordneten Segmente weiterleiteten. Neu war lediglich, dass im Fall, dass es kein höheres Segment gab, ein solchen neu angelegt wurde.
Das sensorische Zentrum des bisherigen cortikalen Segments war mit dem sensorischen Zentrum des neuen Segments über Neuronen der Klasse 4 verbunden. Deren Axone transportierten die Signale der neuen Submodalität aus dem bisherigen cortikalen Divergenzmodul in das übergeordnete sensorische Zentrum des neuen Segments. Ebenso war das motorische Zentrum des cortikalen Segments mit dem motorischen Zentrum des neuen, übergeordneten Segments verbunden.
Und letztlich war in diesem neuen Segment auch das sensorische Zentrum mit dem motorischen Zentrum über Neuronen der Klasse 3 verknüpft. Gleichzeitig projizierte das neue Segment über Neuronen der Klasse 2 in Seitenwechselkerne, die zur Gegenseite projizierten und dort die kontralaterale Hemmung möglich machten.
Auch die Mittelwertzentren empfingen (ab einer gewissen Evolutionsstufe) die Signale dieses neuen Leitersegments und bewirkten eine Rückprojektion zu ihm.
So entstand neben dem primären Cortex ein sekundärer Cortex, der Input von ihm erhielt und zu ihm zurückprojizierte. Wir können diesen sekundären Cortex auch als Assoziationscortex bezeichnen, weil er – erstmalig in der Evolution – keine direkten Rezeptorsignale mehr empfing, sondern die Signale der Submodalitäten aus den Divergenzmodulen (und anderen Modularten) des primären Cortex. Und diese wurden so behandelt, als seine sie Rezeptorsignale.
Wir stellen die schrittweise Entstehung dieses sekundären Cortex in einer Abbildung dar, die sich auf das Leitersystem bezieht. Dargestellt ist, wie immer neue Leitersegmente über dem primären, cortikalen Leitersegment entstanden und sich jeweils seitlich anlagerten, wobei alle Axone nun auf halbkreisförmigen Bahnen innerhalb der weißen Substanz verlaufen mussten.
Rein theoretisch könnte man diese Leitermodule auch klassisch in Form einer einzelnen Leiter anordnen, wäre da nicht die Schädeldecke, die dies verhindert.
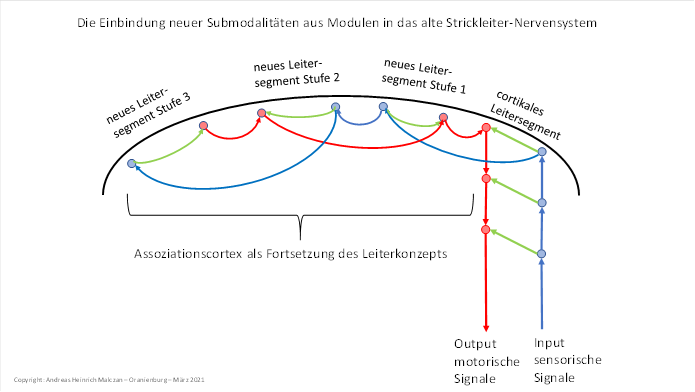
Abbildung 51: Die Einbindung neuer Submodalitäten in das alte Strickleiter-Nervensystem
Eine Folge dieser seitlichen Anlagerung der neu entstandenen Leitersegmente war die seitliche Ausdehnung der Strukturen. Wir bezeichnen sie heute beim Menschen, aber auch bei den Wirbeltieren gemeinhin als Cortex. Wir können diese Sprachregelung beibehalten, sollten uns aber stets ins Bewusstsein rufen, dass der ursprüngliche Cortex das oberste Leitersegment darstellte, welches seinen Input noch über klassische Rezeptoren erhielt.
Nun, auch nach der Auffaltung der obersten, cortikalen Leiter in vier verschiedene Subleitern unterschiedlicher Modalitätengruppen – die Lobi des Gehirns gingen daraus hervor – begann jede dieser Teilleitern ein seitliches Wachstum, um die neu entstehenden, übergeordneten Leitersegmente für die selbsterzeugten Submodalitäten unterbringen zu können.
Wir verzeichnen also ein seitliches cortikales Wachstum nicht nur durch die Signaldivergenz in der Fläche, sondern durch die Entstehung von Submodalitäten, die einen Assoziationscortex entstehen ließen.
Die Entstehung dieses Assoziationscortex seitlich vom bestehenden, primären Cortex erfolgte in allen Lobi des Gehirns und auf beiden Körperseiten. In jedem Gehirnlappen entstanden durch die (recht früh einsetzende) Signaldivergenz die Divergenzmodule, deren Arbeitsweise überall gleich war, deren Input sich jedoch unterschied. Denn bereits recht früh hatten sich die Rezeptoren in Unterklassen zusammengefunden, die jeweils einen künftigen Gehirnlappen für sich beanspruchten.
Inwieweit dieser Assoziationscortex bzw. seine Vorstufe, die Assoziationsleitern, bereits bei den Wirbeltieren der frühen Evolutionszeit entstanden, sei hier nicht weiter diskutiert. Sicherlich entstand der Assoziationscortex des modernen Menschen schrittweise aus dem frühen Strickleiter-Nervensystem.
Dies erklärt auch die Verbindungen der Cortexfelder untereinander. Stets findet man im Assoziationscortex abwechselnd sensorische und motorische Teilgebiete, die untereinander über Neuronen der Klasse 4, der Klasse 3 und der Klasse 5 verknüpft sind. Auch die Mittelwertneuronen der Klasse 6 und 1 – in der obigen Abbildung weggelassen – existieren weiterhin auch in den Assoziationsgebieten. Ebenso projizieren die Gebiete des Assoziationscortex über Neuronen der Klasse 2 in die Gegenseite, deren Projektionsaxone bilden im Gehirn die Struktur des Balkens. Über die Projektion in die dopaminerge Substantia nigra pars compacta und die Rückprojektion zum Striatum sind diese neuen Gehirnstrukturen ebenso in das Basalgangliensystem eingebunden. Weiterhin empfängt das Cerebellum über die Brückenkerne die neuen Elementarsignale, während die Mittelwertprojektion an den Kletterfasern des Pontocerebellums endet. So können aus den Signalen dieser neuen Assoziationsgebiete wieder neue Komplexsignale abgeleitet werden. Auch die Einspeisung in das limbische System erfolgt wie bisher.
Wir verdanken unsere Intelligenz (unter anderem) drei Prinzipien, die sich im Verlauf der Evolution der zentralen Nervensysteme etablieren konnten:
· Möglichst jedes Signal wurde in den Mittelwertkernen berücksichtigt. Dies betraf auch Signale, die erst durch die Arbeit von neuronalen Modulen entstanden waren und somit neue Submodalitäten darstellten.
· Jedes neue Signal, welches in einer Etage des segmentierten Nervensystems durch die Arbeit von neuronalen Modulen entstand, wurde an die übergeordneten Etagen weitergegeben.
· Gab es keine höhere Etage dafür, so begründeten die Projektionsneuronen der Klasse 4 eine neue Etage, so dass ein übergeordnetes neuronales Segment entstand. Dieses musste, bedingt durch die Begrenzung durch die Schädeldecke, seitlich ausweichen. So bildete sich der sekundäre Assoziationscortex.
Möglicherweise konnte das Kopfskelett bei einigen Arten auch in die Höhe wachsen, so dass dort neue Etagen des Assoziationssystems einfach oben angefügt wurden. Dadurch wurde der Kopf kolbenförmig verlängert.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan