Gehirntheorie des Menschen
ISBN 978-3-00-068559-0
Monografie von Dr. rer. nat. Andreas Heinrich Malczan
13 Die drei Subsysteme des menschlichen Gehirns
Die Signalverarbeitung im menschlichen Gehirn und im Wirbeltiergehirn beruht (unter anderem) auf drei Prinzipien, drei Signalklassen und drei Subsystemen.
Prinzip der Divergenzmodule
Durch divergente Verteilung neuronaler Signale und das Wirken elementarer Naturgesetze entstehen im Gehirn neue Signalklassen. Bei der ungedämpften Signalausbreitung können in einem Phasenmodul die Phasenunterschiede erkannt und genutzt werden, um die Schallrichtung zu analysieren.
Bei der gedämpften Signalausbreitung mit nichtlinearer Dämpfung können extremwertcodierte Signale gewonnen werden. Ist die neuronale Übertragungsfunktion dabei streng konkav, so entstehen maximumcodierte Signale. Dies ist die bevorzugte Arbeitsweise der Module in den primären sensorischen Cortexgebieten des menschlichen Gehirns. Die Lage der Extremwerte verschlüsselt die Werte der beteiligten Urgrößen, die ihrerseits den Rezeptoroutput erzeugen. Der Output der Divergenzmodule verschiedener Modalitäten bildet neue Submodalitäten, die im Gehirn neue Etagen (Segmente) bilden.
Prinzip der Transformation und Rücktransformation
Wird im Signalweg zwischen einem Rezeptor und seinem motorischen Ziel durch ein Modul eine Signaltransformation durchgeführt, so muss ein dazu inverses Modul aus den neuen Signalen durch eine Rücktransformation die ursprünglichen Signale möglichst gut rekonstruieren und dem ursprünglichen Signalweg zuführen, während die neuen Signale ein übergeordnetes System in einer neuen Auswertungsschleife bilden.
Prinzip der rückgekoppelten neuronalen Netze im Pontocerebellum
Das Pontocerebellum ist in der Lage, die Aktivität einer cortikalen Neuronenpopulation durch die Aktivität eines einzelnen Neurons zu substituieren. Einer Menge von aktiven Elementarsignalen wird so ein Komplexsignal zugeordnet.
Im inversen Pontocerebellum werden die Komplexsignale wieder in ihre aktiven Elementarsignale zurücktransformiert.
Das Pontocerebellum erkennt das zu einer Neuronenpopulation gehörende Komplexsignal auch dann, wenn nicht alle seine Elementarsignale aktiv sind. Das Komplexsignal hat dann jedoch eine geringere Feuerrate, die mit der Anzahl der aktiven Elementarsignale zunimmt.
Die Rückführung des erkannten Komplexsignals ins inverse Pontocerebellum führt dazu, dass sämtliche ihm zugeordneten Elementarsignale aktiviert werden, auch diejenigen, die zuvor inaktiv waren. Die zusätzlich aktivierten Elementarsignale stellen den selbsterschaffenen Input dar, der im Cortex das Bewusstsein erreicht. Die so aktivierten Elementarsignale können mit anderen, ebenfalls durch Signalvervollständigung erzeugten Elementarsignalen neue Komplexsignale aktivieren. Damit ist das Gehirn fähig, eigene Signale zu erschaffen, die das Bewusstsein erreichen. Diese selbsterzeugten Signale erreichen sowohl die primären als auch die sekundären Cortexgebiete, wo sie uns bewusst werden. Insofern ist das Pontocerebellum der materielle Sitz des Unterbewusstseins. Dies ist die Grundlage unseres Bewusstseins. Es entsteht in uns durch die Fähigkeit, unvollständige Komplexsignale zu erkennen und zu vervollständigen.
Die ständige Signalrotation in diesem rückgekoppelten System, der mit dem ständigen Wechsel der Signalformen zwischen der Elementarform und der Komplexform verbunden ist, bildet die signaltheoretische Grundlage der Intelligenz.
Es gibt im Gehirn der Wirbeltiere und des Menschen generell drei Signalklassen:
· Analoge Signale
· Extremwertcodierte Signale
· Komplexsignale
Analoge Signale werden durch ihre Feuerrate charakterisiert, sie verschlüsselt den Wert einer Größe, die in einem Rezeptor Aktionspotentiale hervorruft. Generell kann die Feuerrate mit der Zunahme der Größe ansteigen, dann sprechen wir vom On-Typ. Dies entspricht in der Mathematik einer streng monoton wachsenden Funktion. Beim Off-Typ nimmt die Feuerrate mit zunehmender Größe ab, die Funktion ist streng monoton fallend. Mischtypen existieren ebenfalls.
Extremwertkodierte Signale entstehen in Divergenzschaltungen mit nichtlinearer Ausbreitungskennlinie. Hier spielt die Krümmung der Übertragungskennlinie eine große Rolle. Bei der räumlichen Ausbreitung in Divergenzmodulen des Gehirns tritt eine nach oben gekrümmte Übertragungskennlinie auf, die auch als streng konkav bezeichnet wird. Sie führt bei der Überlagerung mehrerer Inputquellen zu einem wohldefinierten Maximum, welches eindeutig die Stärke der Urgrößen verschlüsselt.
Die extremwertcodierten Signale können in Konvergenzschaltungen wieder in ihre Ausgangssignale transformiert werden, wobei auch ein gewisser, meist kleiner Signalfehler auftritt.
Analoge und extremwertcodierte Signale können zur Bildung von Komplexsignalen im Pontocerebellum verwendet werden. Im inversen Cerebellum können sie wieder in die Ausgangssignale transformiert werden.
Den drei Signalklassen entsprechen drei Subsysteme im Gehirn.
· Das primäre Subhirn verarbeitet analoge Signale, übergibt sie an das sekundäre Subsystem und empfängt vom sekundären System die analogen Signale, die durch Konvergenzmodule aus den extremwertcodierten Signalen gewonnen werden.
· Das sekundäre Subhirn empfängt vom primären Subhirn die analogen Signale und unterwirft sie einer Signaldivergenz, wodurch extremwertcodierte Signale entstehen. Diese werden an das tertiäre Subhirn weitergegeben. Auf der absteigenden Seite empfängt das sekundäre Subhirn die extremwertcodierten Signale des inversen Pontocerebellums.
· Das tertiäre Subhirn verarbeitet Elementarsignale, die zu analogen oder extremwertcodierten Signalen gehören, nachdem diese einer lateralen Nachbarhemmung unterzogen wurden und somit spärlich codiert sind. Es ordnet den aktiven Neuronengruppen ein Komplexsignal zu, dies erfolgt im Pontocerebellum. Auf der absteigenden Seite werden aus den Komplexsignalen wieder die ursprünglichen Elementarsignale gewonnen und an das sekundäre System übergeben.
Jede dieser Substrukturen ist in der Lage, ihren Output in die limbischen Rotationsschleifen einzuspeisen, so dass sie dort temporär aktiv sind und mit Gegenwartssignalen kombiniert werden können. Wir bezeichnen diese Speicherart als Rotationsspeicher. Über den Nucleus anterior des Thalamus stehen die limbischen Signale dem System wieder zur Verfügung.
Ebenso kann jedes dieser drei Subsysteme seinen Output in die Basalganglien einspeisen, die eine hemmende und zeitverzögerte Kopie dieser Signale erzeugen, die mit Gegenwartssignalen überlagert wird, um Signalveränderungen und Bewegungen in allen Modalitäten zu erkennen. Ebenso wird eine erregende und zeitverzögerte Kopie angefertigt. Dies geschieht mit der Substantia nigra pars reticularis als letzter Station der Basalganglien. Die erregende und zeitverzögerte Kopie erreicht das Pontocerebellum und kann mit Gegenwartssignalen kombiniert werden zu neuen Komplexsignalen.
Durch die ständige Signalweitergabe von den unteren zu den oberen Systemen auf der aufsteigenden Seite und die Weitergabe der Signale von oben nach unten in der absteigenden Seite stehen alle drei Signalklassen zeitgleich zur Verfügung. Da der Output eines Systems gleichzeitig der Input des benachbarten Systems ist, kommt es zu einer Signalrotation.
Komplexsignale sind gekennzeichnet durch die Aktivität eines einzelnen Neurons. Daher können sie in das primäre System eingespeist werden, wo sie den analogen Signalen gleichgestellt werden und als neuer Input weitere Analyseresultate hervorrufen.
Prinzip der Signalrotation
Während im aufsteigenden System Signale durch Transformation in Modulen in die übergeordnete Signalklasse transformiert werden, erfolgte im absteigenden System die Rücktransformation dieser Signale in die ursprüngliche Signalklasse, sie werden dem ursprünglichen System als Input zugeführt. Dadurch kommt es im System zur ständigen Signalrotation mit ständigem Wechsel der Signalklassen. (Oszillationstheorem von Malczan, frühere Monografie)
Während der Signaloszillation im System werden die unvollständig erkannten Komplexsignale ergänzt und vervollständigt, indem die fehlenden Elementarsignale im inversen Cerebellum aktiviert werden. So erzeugt das System Eigeninput, der dem inversen Cerebellum entsteigt und im Cortex ankommt, wo er uns bewusst wird.
Prinzip der Ergänzung von unvollständig erkannten Komplexsignalen
Komplexsignale werden auch als solche erkannt, wenn mindestens die Hälfte der zum Signal gehörenden Elementarsignale aktiv sind. Das Erkennungssignal ist ein Komplexsignal, welches im absteigenden System in seine Elementarsignale transformiert wird. Dabei werden alle Komponenten der ihm zugeordneten Elementarsignale aktiviert, auch die vorher inaktiven. Dadurch findet im Gehirnsystem ständig eine Signalergänzung statt, bei der die nichtaktiven Signalanteile eines unvollständigen, aber erkannten Signals aktiviert werden, indem das aktive Komplexsignal im absteigenden System wieder alle zu ihm gehörenden Elementarsignale aktiviert. Diese im absteigenden Signalweg aktivierten Elementarsignale, die zuvor inaktiv waren, stellen die vom System selbsterschaffenen Ergänzungssignale dar. Sie entsteigen dem Unterbewusstsein, indem sie mit anderen, ebenfalls durch Signalergänzung induzierten Elementarsignalen im Cerebellum ihrerseits neue Komplexsignale aktivieren, deren Bestandteile sie in der Elementarform darstellen. Dies ist das Prinzip der Entstehung von Gedanken, Gefühlen, Anschauungen, Vorstellungen und der Grundalgorithmus jeglicher Intelligenz.
Das dreistufige Gehirnsystem der Wirbeltiere und des Menschen ist nachfolgend dargestellt.
Ich gehe davon aus, dass menschliche Intelligenz genau von diesem System hervorgebracht wird.
Der wichtigste Unterschied zwischen dem Gehirn des Menschen und dem Gehirn anderer Wirbeltiere besteht darin, dass er in der Lage war, dem ursprünglichen Strickleitersystem der Wirbeltiere zusätzliche Assoziationsleitern hinzuzufügen.
Die Entwicklung der Sprache erlaubte es ihm, eine Komprimierung der Signale vorzunehmen.
Man vergleiche als Beispiel die zur Abspeicherung eines kompletten Films nötigen Speicherplatz (bei dem eine Datenkomprimierung nicht angewendet werden darf) mit dem Speicherplatz, den das zum Film gehörende Drehbuch benötigt. Dann wird schnell klar, dass die Datenkomprimierung durch die Erfindung der Sprache zur Einsparung von neuronalem Speicherplatz führt.
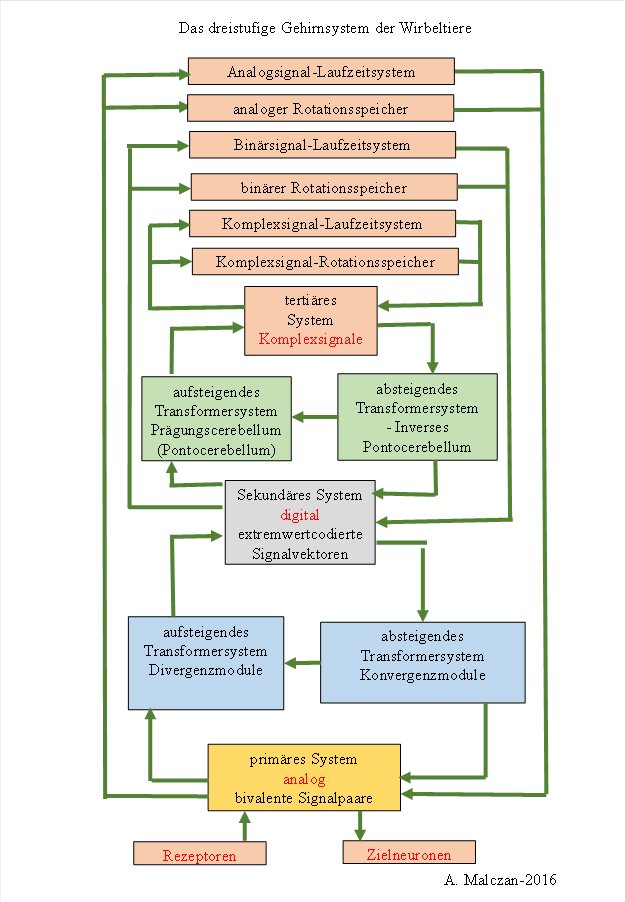
Abbildung 52: Die drei Subsysteme des Gehirns: primäres, sekundäres und tertiäres System
Monografie von Dr. rer. nat. Andreas Heinrich Malczan