Gehirntheorie des Menschen
ISBN 978-3-00-068559-0
Monografie von Dr. rer. nat. Andreas Heinrich Malczan
2. Die Aufspaltung der Modalitäten und die Modulentstehung – Voraussetzung für die Arbeitsteilung im Gehirn
Die Aufspaltung der Modalitäten begann schleichend und vollzog sich schrittweise. Unterschiedliche Rezeptoren lieferten Signale unterschiedlicher Modalitäten. Bereits bei der Ankunft der Signale im Neuralrohr, wo sie an Neuronen der Klasse 3 und 4 endeten, kam es zur Trennung. Unterschiedliche Rezeptorarten beanspruchten eigene, nur ihnen zugeordnete Neuronenschichten im Neuralrohr. Hierbei war der räumliche Zusammenhalt der Neuronen einer Modalität so stark, dass sie segmentübergreifend war. Neuronen der gleichen Modalität fanden sich im Neuralrohr auch dann in einer Schicht zusammen, wenn ihnen Signale aus unterschiedlichen Segmenten entsprachen. Dann waren sie innerhalb der Schicht räumlich benachbart, wenn die Segmente benachbart waren.
Dabei bildete sich eine topologische Wohlordnung heraus. Signale von benachbarten Rezeptoren der gleichen Modalität endeten im Neuralrohr in benachbarten Neuronen. Da sensorische Signale – und alle Rezeptorsignale waren ja sensorischer Art – aus allen Segmenten kopfwärts zogen, besaß jedes Kopfsegment eine komplette Kopie aller Rezeptorsignale der Rumpfsegmente. Hierbei wurde die Anordnung der Rezeptoren in der Körperoberfläche (Haut) oder im Körper von den Rezeptoren auf die Eingangsneuronen des Körpermodells im jeweiligen Kopfsegment übertragen.
So besaß jedes Kopfsegment ein komplettes Körpermodell der Rumpfrezeptoren, das topologieerhaltend war.
Das oberste Kopfsegment entspricht beim Wirbeltier dem Cortex. Auch hier endeten die Rumpfsignale topologisch wohlgeordnet und spiegelten die Anordnung der Rezeptoren im Körper des Tieres wieder. Diese topologische Anordnung wird auch als Homunculus bezeichnet. Sie ist im Cortex jedoch mehrfach vorhanden. Offenbar trennen sich die kopfwärts ziehenden Axone, welche die Signale verschiedener Modalitäten transportieren, vor dem Erreichen des ersten Segments auf, jede Modalität zieht in ein eigenständiges Cortexgebiet.
Dies bezeichnen wir als Aufspaltung der Modalitäten. Sie begann einst damit, dass sich bereits beim Strickleiternervensystem eine Dreiteilung der Rezeptorarten ausbildete, so dass sich die Strickleiter an den oberen zwei Segmenten aufteilte. Statt einer Leiter gab ab dem zweiten Segment drei Leitern, jede für eine bestimmte Modalitätengruppe.
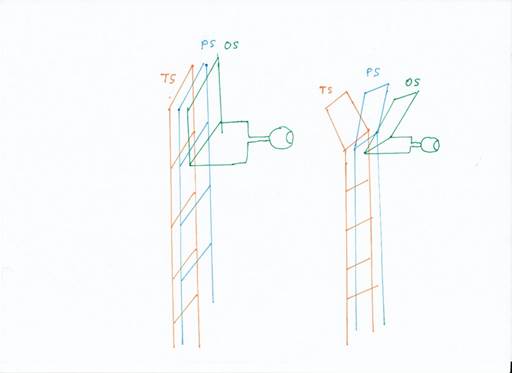
Abbildung 4: Die Aufspaltung der Modalitäten
Da die Signale in den sensorischen Holmen aufwärts zogen, dann in einer Art Schleife die Richtung umkehrten und wieder abstiegen, bezeichnen wir die oberste Etage auch als Schleife (oder Wendeschleife). Dann gab es nach der Aufspaltung der urtümlichen Modalitäten drei Schleifen: die Temporalschleife, die Parietalschleife und die Occipitalschleife.
Aus diesen drei Leitern gingen drei Cortexlappen der Wirbeltiere hervor, der Parietallappen, der Temporallappen und der Occipitallappen. Der Parietallappen empfing die Signale der Rezeptoren, die direkt mit der Motorik zusammenhingen, etwa die Muskelspannungsrezeptoren, die Rezeptoren für taktile Reize und Schmerzreize. Der Temporallappen nahm die Rezeptorsignale der Haarzellen aus dem Vestibularsystem entgegen, später die Seitenliniensignale und die Hörsignale. Der Occipitallappen empfing visuelle Signale.
Mit der Ausbildung des Cerebellums entstand der Frontallappen, der den Output des Cerebellums empfängt.
Dies sind die vier wichtigsten Lobi des Gehirns.
Während der evolutionären Höherentwicklung der Wirbeltierarten schritt die Aufspaltung der Modalitäten weiter voran. Nun trennten sich die Modalitäten nicht nur auf der Höhe des zweiten Segments, dessen sensorische und motorische Kerne nun als Thalamus bezeichnet werden, sondern bereits in tieferen Segmenten.
Die Trennung ging schließlich – etwa bei den Primaten und beim Menschen – so weit, dass bestimmte Muskelgruppen eigene Eingangskerne in den Kopfsegmenten bildeten, die eine eigene Repräsentation im Cortex erhielten. Dies war für die Interaktion von Signalen verschiedener Modalitäten ein enormer Fortschritt, denn nun konnten beispielsweise Augenbewegungen, Halsbewegungen und Bewegungen visueller Objekte miteinander so koordiniert werden, dass ein stabiles Bild entstand, unabhängig von eigenen oder fremden Bewegungen. Dies ging nur, weil zuvor eine saubere Trennung der Modalitäten erfolgte.
Die Aufspaltung der Modalitäten erreichte beim Menschen einen besonders hohen Ausprägungsgrad. Entstanden zu einer Modalität neue Submodalitäten, so trat in den Eingangsschichten der zweiten und ersten Etage, also im Thalamus und im Cortex, eine Trennung dieser Submodalitäten in separate Schichten auf.
Diese Auftrennung in submodale Schichten begann natürlich nicht erst beim Menschen, sondern viel früher, wahrscheinlich bereits bei einfachen Wirbeltieren. Sie erreichte jedoch beim Menschen einen so hohen Ausprägungsgrad, dass sein Gehirn Leistungen erbringen kann, die im Tierreich entweder unmöglich sind oder nur bei wenigen Spezies – und dann auch immer nur selektiv als Spezialfall – auftreten. So können manche Vogelarten kognitive Leistungen erbringen, zu denen die übrige Tierwelt nicht fähig ist, gleichwohl jedoch der Mensch.
Die Aufspaltung in Submodalitäten soll beispielhaft erklärt werden.
Ursprünglich gab es zur visuellen Wahrnehmung nur eine Rezeptorart: den Grünrezeptor. Er versorgte die Inputschicht 4 mit Signalen, die die aktuelle Helligkeit bewerteten. Der Output wurde an die benachbarten Schichten 3 und 2 und 6 übertragen. Die Schicht 3 projizierte zum motorischen Zentrum, die Schicht 2 zum Seitenwechselkern zwecks kontralateraler Hemmung und die Schicht 6 versorgte die Mittelwertkerne. In der folgenden Abbildung ist dies als Stufe 1 interpretiert.
Durch Genverdopplung bildete sich ein neuer Rezeptor: der Rotrezeptor. Nun konnten zwei Farben wahrgenommen werden.
Dies führte im sensorischen Kern des zweiten Segments – dem visuellen Thalamus – zu einer Aufspaltung der vierten Eingangsschicht. Dort empfingen die Neuronen den Input von der Retina. Die Schicht 4 spaltete sich auf in die Subschichten S4-grün und S4-rot. Hierbei – so die Hypothese, besaß jede der Subschichten eine eigene Outputschicht aus Neuronen der Klasse 3.
Über Neuronen der Klasse 3 zog dieser Input zum motorischen Teil des visuellen Thalamus und endete dort an Neuronen der Klasse 5, die zu den Motoneuronen projizierten. So konnte das Tier auf visuelle Reize mit Bewegungen reagieren, die eine Annäherung an Futter oder die Flucht vor Fressfeinden bewirkten.
Über Neuronen der Klasse 2 zogen die Signale zur Gegenseite, um dort kontralaterale visuelle Reize zu hemmen. So standen beide Augen in neuronaler Konkurrenz, das stärkere Signal unterdrückte das schwächere und setzte sich durch. Größere Beute auf der linken Seite wurde bevorzugt gefressen, wenn die Beute rechts deutlich kleiner war.
Über Neuronen der Klasse 6 wurden die visuellen Mittelwertkerne aktiviert und kurbelten über Rückprojektionen in sensorische und motorische Zentren die Futtersuche an.
Als sich nun der Grünrezeptor aufspaltete und dadurch ein neuer Rotrezeptor entstand, führte dies im Thalamus und im Cortex ebenfalls zur Aufspaltung der vierten Inputschicht.
Sie zerfiel in zwei Subschichten. Die Schicht 4, die bereits seit Urzeiten den Input vom Grünrezeptor erhielt, blieb erhalten. Wir geben ihr jedoch eine neue Bezeichnung, sie ist nun die Schicht 4-grün.
Die neuen Rotrezeptoren sandten ihre Signale auch in die Schicht 4. Anfänglich mögen die grünen und die roten Signale in der Schicht 4 bunt gemischt gewesen sein. Doch Gleich und Gleich gesellt sich gern! So begannen die Inputneuronen, die die roten Signale empfingen, nach oben zu wandern, und bildeten dann (nach tausenden oder hunderttausenden von Generationen) eine eigene Schicht 4-rot. Noch heute ist in der Neurologie die Ansicht weit verbreitet, dass sich neue (evolutionär jüngere) Neuronen immer außen an die vorhandenen anlagern und eine eigene Schicht bilden.
Damit war die Trennung der zwei Submodalitäten im visuellen Thalamus vollzogen. Sie übertrug sich jedoch im Verlauf der Evolution auch auf das übergeordnete Segment. So trennten sich die vom Thalamus über die Neuronen der Klasse 4-rot und 4-grün aufsteigenden Signale in der vierten Cortexschicht ebenfalls auf. Die vierte Cortexschicht zerfiel in die Subschichten 4-grün und 4-rot. Auch hier war die evolutionär neuere Schicht – die Schicht 4-rot – oben über der älteren Schicht 4-grün angeordnet. In der folgenden Abbildung ist dies als Stufe 2 gekennzeichnet.
Als noch später die blauempfindlichen Rezeptoren in der Retina entstanden, standen drei Rezeptorarten für die Schicht 4 im Thalamus und im Cortex zur Verfügung. Daher musste eine neue Schicht 4-blau entstehen, die sich räumlich wieder ganz oben anordnete, weil sie die jüngste Schicht war.
Ursprünglich gab es zur Schicht 4 die darüberliegende Schicht 3. Als die Aufspaltung der Modalitäten begann, wurde auch diese Schicht 3 in die Aufspaltung einbezogen. Jede neue Subschicht 4 erhielt ihre eigene Subschicht 3. Dies ist in der unteren Abbildung bereits berücksichtigt. Dies traf wahrscheinlich auf alle sechs Neuronenschichten zu, die nun eigene Subschichten bildeten.
Beim Menschen jedoch – und möglicherweise schon bei früheren Wirbeltieren – ordnete sich die Schicht 4-blau auch (zusätzlich) unterhalb der bisherigen Schichten an. Ein Grund war (möglicherweise) die Signalverwandtschaft zwischen dem grünempfindlichen und dem blauempfindlichen Rezeptor. Letzterer war quasi der „Enkel“. So gab es zwei Blau-Schichten, eine ganz oben und eine ganz unten. Dies verdeutlicht die nachfolgende Abbildung in der Stufe 3.
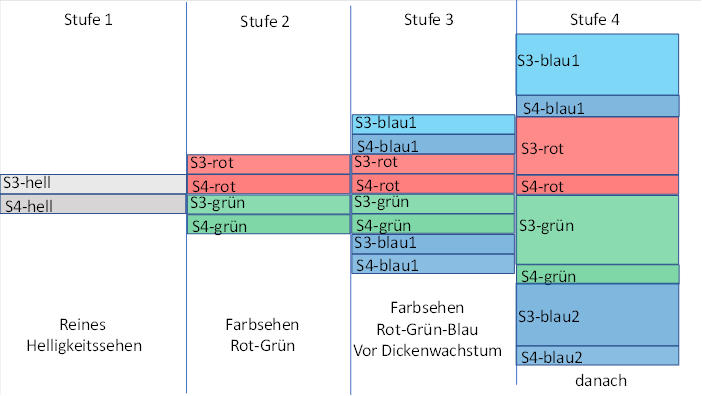
Abbildung 5: Aufspaltung der visuellen Modalitäten
Der Vorteil lag auf der Hand. Nun konnte man sowohl die Einzelfarben Grün, Rot und Blau sehen als auch Mischfarben, etwa Rot + Blau = Violett, Rot + Grün = Gelb oder Grün + Blau = Grünblau. Dann waren die Outputneuronen der jeweils zwei benachbarten Schichten gleichzeitig aktiv. Wir nennen dies additive Farbmischung. Die fand nun auch neuronal statt. Ein einfaches Rechenbeispiel soll dies verdeutlichen.
Ein Rot-Rezeptor der Retina möge eine Ganglienzelle erregen, deren Feuerrate mit einer Frequenz von 100 Aktionspotentialen pro Sekunde ein Inputneuron im visuellen Cortex erreicht. Dieses befindet sich in der Schicht S4-rot. Seine Erregung aktiviert ein Outputneuron der Schicht S3-rot. Bei der extrem kurzen Entfernung – beide Schichten sind direkt benachbart – geht die Erregung komplett auf das Outputneuron über, dieses feuert dann auch mit 100 Hz.
Wenn nun stattdessen ein Grün-Rezeptor ebenso stark feuert, weil grünes Licht ihn erregt, so aktiviert er ein Outputneuron in der Schicht S3-grün. Dieses feuert nun ebenfalls mit 100 Hz.
Wir stellen uns nun vor, beide Rezeptorarten wären erregt, weil gelbes Licht auf die Retina fällt. Gelbes Licht erregt jedoch den Rot-Rezeptor, wie man unschwer im Kennliniendiagramm der retinalen Empfindlichkeitskurven der verschiedenen Sehpigmente ersehen kann. Etwas weniger als rotes Licht es tun würde. So feuert die Rot-Ganglienzelle beispielsweise nur mit 80 Hz.
Gleichzeitig feuert die grünempfindliche Gangleinzelle, denn auch das grüne Sehpigment reagiert auf Gelb, wenn auch schwächer als auf Grün. So feuert auch in der Schicht S3-grün ein Outputneuron mit beispielsweise 75 Hz.
Da der Rotrezeptor aus dem Grünrezeptor hervorging, waren die Neuronen für den Signalweg der roten und der grünen Farbe signalverwandt. Daher konnten die Outputneuronen der Schicht S3-rot auch die Erregung von Inputneuronen der Schicht S4-grün empfangen. So bildete sich dort eine Untergruppe von Neuronen, die beide Signalarten empfangen konnten.
So kam es – im Verlauf der Evolution – dazu, dass bei gleichzeitiger Aktivität eines Inputneurons in der Schicht S4-grün und in der Schicht S4-rot ein gemeinsames Outputneuron in der Schicht S3-grün kontaktiert wurde und ebenfalls erregt wurde. Wir wollen dieses Outoutneuron vorläufig als Additionsneuron bezeichnen. Zusätzlich zu den bereits erregten in der Schicht S3-grün und S3-rot. Diese empfingen jedoch nur Input von einem Neuron, feuerten also mit 80 Hz für Grün und 75 Hz für Rot. Nun gab es eines, welches Input von beiden bekam. Seine Feuerrate war größer als 80 und auch größer als 75 Hz, beispielsweise 155 = 80 + 75.
Damit gibt es für jedes Bildpixel der Netzhaut drei Outputneuronen: Eines ist für die rote und eines für die grüne Farbe zuständig, das dritte wurde additiv versorgt, empfing also gleichzeitig den Input für die rote und für die grüne Farbe. Waren sowohl der Rotrezeptor als auch der Grünrezeptor erregt, so war der Output des Additionsneurons deutlich stärker als der Output der zwei Neuronen für Rot und Grün.
Hier kommt die Nachbarhemmung ins Spiel. Die Feuerrate von 155 Hz ist deutlich stärker als die von 80 Hz oder 75 Hz. Sie hemmt diese beiden Feuerraten bzw. die sie erzeugenden Neuronen. Bei totaler Hemmung gibt es nur einen Output: f = 155 Hz. Und er signalisiert eindeutig: Gelbes Licht erkannt.
Denn der übrige Output des Systems ist vernachlässigbar schwach, weil er ein Opfer der gegenseitigen Hemmung der Outputneuronen ist.
So lernte der visuelle Cortex, die Farbe Gelb zu erkennen. Den gleichen Gedankengang kann man für die Mischfarben Violett = Rot + Blau oder für Blaugrün = Blau + Grün anstellen. Dieses System konnte Farben erkennen, die sich aus zwei der drei Grundfarben Rot, Grün und Blau additiv zusammensetzte.
Es muss darauf hingewiesen werden, dass die Aufspaltung der visuellen Modalitäten nicht erst im menschlichen Cortex nachweisbar ist. So tritt sie bereits in der Retina auf. Dort enden die Axone der verschiedenen Rezeptoren in unterschiedlichen Schichten der Retina. Und bereits in der Retina existieren Neuronen – man nennt sie dort Ganglienzellen – welche Input aus unterschiedlichen Schichten beziehen. Dieser Input kann erregend, aber ebenso auch hemmend sein. So gibt es eine Art von Ganglienzellen, die vom Grün-Rezeptor erregt, vom Rot-Rezeptor jedoch gehemmt werden. Wir bezeichnen sie als Grün-On/Rot-Off-Ganglienzellen. Ebenso gibt es eine Art von Ganglienzellen, die vom Rot-Rezeptor erregt, jedoch vom Grün-Rezeptor gehemmt werden. Sie werden als Grün-Off/Rot-On-Ganglienzellen bezeichnet. Eine weitere Ganglienzellenart ist vom Typ Blau-On/Gelb-Off. Weiterhin gibt es – im Prinzip als Neuronenklasse 6 – große Mittelwertneuronen, die sämtliche On-Ganglienzellen anzapfen und den Typ Hell-On darstellen. Ebenso gibt es den magnocellularen Dunkel-On-Typ. Insofern stellt die Retina bereits ein Modul dar. Sie liefert bereits helligkeitsbewertete und farbbewertete Signale an das Tectum opticum, den visuellen Thalamus und den Cortex. Die Schichtung in der Retina setzt sich im Thalamus und im Cortex fort und wird noch weiter verfeinert.
Beim Menschen – so meine Theorie – ging die Aufspaltung der Modalitäten im Farbsystem weiter. Während im primären visuellen Cortex die Signale der drei verschiedenen Farbrezeptoren noch in verschiedenen Subschichten der Schicht 4 endeten, entstand über der bisherigen cortikalen Etage ein neues Subsystem, eine Art neue Etage des Strickleitersystems. Dort endeten die Farbinformationen nicht mehr in drei verschiedenen Inputschichten, sondern in einer einzigen Schicht. Dafür spalteten sich die Signale jedoch auf und zogen alternierend in diese neue Etage. So kann es, dass dort, im sekundären visuellen Cortex, ein neues sensorisches Inputgebiet entstand. In diesem Inputgebiet war die Retina wiederum retinotop abgebildet. Jedem Retinapixel entsprachen hier jedoch drei Inputneuronen, die ihrerseite ein Dreieck bildeten. In einer Ecke des Dreiecks befand sich das Inputneuron, welches vom primären Cortex das Signal Rot-On/Grün-Off empfing. In der zweiten Ecke des Dreiecks war das Inputneuron positioniert, welches das Signal Grün-On/Rot-Off des zugehörigen Retinapixels erhielt. In der dritten Ecke des Dreiecks traf das Blau-Signal ein, welches jedoch von der Farbe Gelb gehemmt wurde und daher das Signal Blau-On/Gelb-Off darstellte. So war jedem einzelnen Retinapixel ein Farbdreieck zugeordnet, dessen Ecken die Grundfarben Rot, Grün und Blau zugeordnet waren. In diesem Cortexgebiet entsprach die Retina in etwa der Fläche eines Farbfernsehers, bei dem jeder Bildpunkt aus drei Lichtpunkten besteht, denen die Grundfarben Rot-Grün-Blau zugeordnet sind. Durch die additive Farbmischung entsteht für uns ein farbiges Abbild der Welt. Und genau so ist das sekundäre visuelle Farbfeld organisiert.
Auch dieses sekundäre visuelle Farbenfeld projizierte (unter anderem) in die Basalganglien und in das limbische System, zusätzlich entstand ein tertiäres Cortexgebiet, das sowohl den Input aus den sekundären Farbfeld empfing als auch den zeitverzögerten Rückkehroutput der Basalganglien, so dass dort Veränderungen und Bewegungen wahrnehmbar wurden. Der Rückkehroutput des limbischen Systems führte dort direkt zu Falschfarben-Nachbildern in Komplementärfarben.
Eine derartige Aufspaltung beobachtet man auch zwischen Thalamus und dem primären Cortex. Im visuellen Thalamus enden die Signal aus dem linken und den rechten Auge noch in verschiedenen Schichten, die aber retinotop wohlgeordnet sind. Diese Signale werden im visuellen Cortex jedoch nicht mehr in übereinander liegenden Schichten angeordnet, sondern bilden alternierend Augendominanzsäulen, deren Input abwechselnd vom linken und vom rechten Auge kommen. Anstelle der Schichtung tritt die alternierende Anordnung auf. Und für das Farbsystem postuliere ich ebenfalls einen Übergang vom Schichtensystem im primären visuellen Cortex zum sekundären Cortex, wo die zuvor in drei Schichten angeordneten Farbsignale nun in der Fläche alternierend angeordnet sind und so die Farbdreiecke bilden.
Diese Entwicklung stellte jedoch nicht den Endpunkt dar. Das Farbsystem schaffte es durch eine kleine Veränderung, Millionen von Farben zu erkennen. Dies schafften jedoch nicht alle Spezies. Für das exzellente Farbsehen war eine wichtige, jedoch relativ einfache Veränderung im Aufbau der Outputschichten der Klasse 3 erforderlich: das Dickenwachstum und das Breitenwachstum. Wir diskutieren hier zunächst das Dickenwachstum. Dies ist in der obigen Darstellung als Stufe 4 dargestellt. In meiner früheren Monografie habe ich die These aufgestellt, dass dieses Dickenwachstum vorwiegend bei Reptilien und Vögeln auftrat. Bei Reptilien (und Vögeln) bildete sich so der dorsale ventrikulärer Kamm, englisch dorsal ventricular ridge (DVR) genannt. Bei Vögeln tritt diese Struktur auch an die Gehirnoberfläche auf und wird dort als Hyperpallium bezeichnet. Beide Strukturen sind offenbar vertikale Divergenzmodule.
Bei Wirbeltieren mag die Entwicklung auch mit dem Dickenwachstum der begonnen haben, es kam jedoch – bei etlichen Spezies – ein Flächenwachstum oder bei Primaten gar ein Dicken- und Flächenwachstum hinzu.
Wie kam es zunächst zum Dickenwachstum?
Jede Outputschicht der Neuronenklasse 3 begann, Reserveneuronen auszubilden. Neuronen konnten absterben. Gab es Ersatzneuronen in der Nähe, konnten diese als Ersatz einspringen.
So wurde es (bei höheren Spezies) zum Standard, dass in den Outputschichten der Klasse 3, aber auch der Klasse 2, 5 und 6 mehr Neuronen gebildet wurden, als unbedingt nötig waren.
Hier entsteht für Neurologen ein Problem: Neuronen mit identischen Signalen hemmen sich gegenseitig immer, wenn sie aktiv sind. Dies führt zur Atrophie der überzähligen Neuronen, so dass nur eines übrigbleibt.
Diese Atrophie tritt jedoch nicht auf, wenn die Neuronen sich in der Aktivität unterscheiden, also genau genommen nicht identische, sondern nur ähnliche Signale verarbeiten.
Zwei Outputneuronen, die von einem oder von mehreren Neuronen mit Input versorgt werden, bilden genau dann etwas unterschiedliche Signale, wenn sich die Wegstrecken der Signale zu ihnen unterscheiden. Wenn ein Neuron näher an den Inputneuronen angeordnet ist, während das zweite weiter entfernt ist, so unterscheiden sich ihre Erregungen voneinander. Denn in myelinfreien Schichten wirkt eine Dämpfung. Je größer die Entfernung, umso größer die Dämpfung. Wenn es günstig war, die Signale über mehrere Signalwege zu übertragen, so mussten die beteiligten Neuronen sich wenigstens durch die Signaldämpfung unterschieden. Denn dann hatten sie unterschiedliche Feuerraten. Ihre Signale waren nur noch ähnlich, jedoch nicht identisch. So fand keine Atrophie statt.
Daher wuchs die Dicke der Outputschichten. So waren genug Reserveneuronen vorhanden. Sie unterschieden sich im Outputverhalten geringfügig, jedoch hinreichend. Denn der Abstand eines Outputneurons zur oberen und zur unteren Inputschicht unterschied sich von jedem Outputneuron, welches darüber oder darunter angeordnet war. Alle zu einem Retinapunkt gehörenden Outputneuronen in den visuellen Outputschichten mit größerer Dicke bildeten eine Augendominanzsäule. Sie empfingen den Input aus der unteren und der oberen Schicht gleichzeitig. Jedes Outputneuron wurde gleichzeitig von der oberen als auch von der unteren Schicht versorgt, die synaptischen Verbildungen dazu wurden bereits beim Herausbilden und beim Ausreifen der Outputschicht angelegt.
Wie konnte dieses System jedoch Farben, insbesondere Mischfarben erkennen?
Hier müssen wir etwas Mathematik betreiben und das Wirkprinzip der Signalmischung erläutern. Wir bezeichnen solche Systeme hier als Divergenzmodule mit vertikaler Signalmischung. Denn die Erregungen der Farbsensoren gelangen zu den zwei Inputschichten – eine unten und eine oben, und breiten sich vertikal aus, um sich zu mischen.
Das Divergenzmodul mit vertikaler Signalmischung wird nachfolgend beschrieben. Es sei bereits hier darauf hingewiesen, dass etliche Spezies der Wirbeltiere dieses Divergenzmodul beträchtlich erweitern konnten, indem sie auch ein Breitenwachstum nutzten. Solche Divergenzmodule ermöglichen im Gehirn die Entstehung von Orientierungssäulen für die visuelle Richtungs- und Konturenanalyse.
Nachdem die Analyseeigenschaften des visuellen Divergenzmoduls mit vertikaler Signalmischung für den parvocellularen Input erklärt wurden, folgt die mathematische Herleitung seiner bemerkenswerten Eigenschaften.
Ein Hinweis sollte hier noch gegeben werden: Die Nachbarhemmung in solchen Divergezmodulen erfolgt erst, wenn der Output das Divergenzmodul verlässt. Der Output der verschiedensten Subschichten der Klasse-3-Neuronen verlässt das System auf mehreren Wegen. Nach oben werden Outputneuronen der Klasse 2 aktiviert, die ihren Input an die kontralaterale Seite weiterleiten. In dieser Schicht muss die Nachbarhemmung stattfinden.
Weiterhin projiziert die Schicht 3 zur motorischen Seite des visuellen Cortex. Zu erkennen daran, dass der sensorische Input über Axone zur motorischen Seite zieht und dort Neuronen der Klasse 5 aktiviert. Die Nachbarhemmung kann nun zwischen diesem Output wirken. Die motorische Seite des visuellen Cortex wird oft als sekundärer Cortex bezeichnet.
Die Nachbarhemmung ist also aus den Divergenzmodulen ausgelagert. Daher werden Neurologen, die mit einer feinen Sonde im Feld V1 die Erregung analysieren, oft den Eindruck haben, als seine alle Neuronen in einer Augendominanzsäule aktiv. Prinzipiell ist dies so, aber dennoch ist ihre Erregung unterschiedlich stark. In den Outputschichten findet eine Extremwertcodierung statt. Das maximal erregte Neuron bewertet die jeweilige Submodalität hinsichtlich ihrer Signalstärke. Seine Aktivität ruft das hervor, was Neurologen und Philosophen als Qualia hinsichtlich eines elementaren Reizes bezeichnen.
Die Aufspaltung der Modalitäten ging jedoch weiter. Gab es ein übergeordnetes Segment, so wurden Signale, die zuvor verschiedene Schichten eines Moduls erreicht hatten, im darüber befindlichen Segment in einer Ebene nebeneinander angeordnet, wobei sich die vorhandenen Submodalitäten naturgemäß vermischen mussten. Gab es jedoch kein höheres Segment, so erschufen diese Signale ein neues Segment, in dem dieser letzte Aufspaltungsschritt vollzogen wurde. Daher gibt es über dem primären visuellen Cortex, der seine Signale noch für jede Submodalität in einer eigenen Schicht sortiert anordnet, einen übergeordneten visuellen Cortex (zweiten Grades), in dem beispielsweise die Farbinformationen der drei Grundfarben Rot, Grün und Blau in einer einzigen Schicht angeordnet sind, so dass dort praktisch kleine Farbdreiecke entstehen. Im Zuge der Signaldivergenz in dieser Region dehnten sich die Farbdreiecke aus und die Outputneuronen dieser Dreiecke konnten wiederum eine Maximumcodierung der Mischfarbe vornehmen, die durch Überlagerung der Erregungen der drei Inputneuronen eines jeden Farbdreiecks realisiert wurde. Erst dadurch konnte eine weitere und viel feinere Farbanalyse vorgenommen werden.
Diese weitere Aufspaltung der Submodalitäten schritt im motorischen Bereich ebenso voran wie im visuellen. Alle Modalitäten dürften davon betroffen sein.
- ← 1. Die Entstehungsgeschichte des Gehirns - das Konzept der Segmente und Neuronenklassen
- 3. Der Wandel der Abbildungstopologie und der Übergang von zylindrischen zu sphärischen Körpermodellen →
Monografie von Dr. rer. nat. Andreas Heinrich Malczan