Gehirntheorie des Menschen
ISBN 978-3-00-068559-0
Monografie von Dr. rer. nat. Andreas Heinrich Malczan
4.6. Das sensorische Schwerpunktmodul
Schwerpunktmodule dienen der Steuerung von Augenbewegungen, der Steuerung der Kopfdrehung sowie der Koordinierung von Rumpfbewegungen unter anderem durch visuelle Signale.
Wir werden dies am Beispiel eines visuellen Schwerpunktmoduls zur Steuerung von Augenbewegungen zeigen. Schwerpunktmodule dienen dem Ziel, ein visuelles Objekt so mit den Augen zu fixieren, dass es im Zentrum des Sehfeldes positioniert wird. Bewegungen des Objekts lösen Augenbewegungen aus, so dass der Blick dem Objekt folgt. Reichen Augenbewegungen allein nicht aus, so erfolgt zusätzlich eine Kopfdrehung oder gar eine Rumpfbewegung. Gesteuert wird dies über Schwerpunktmodule.
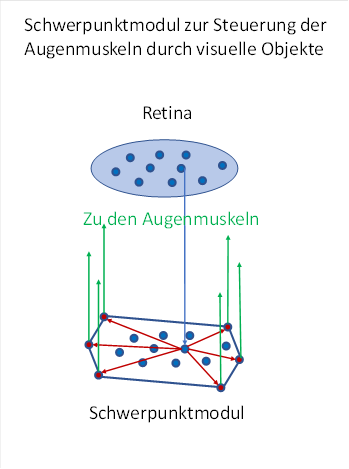
Abbildung 36: Schwerpunktmodul - vereinfachte Darstellung
In der obigen Darstellung sind in der Retina die magnocellularen Ganglienzellen symbolisch dargestellt. Jede magnocellulare Ganglienzelle ist mit einem Neuron im Schwerpunktmodul verbunden, wobei die Anordnung der Ganglienzellen auf die Schwerpunktneuronen retinotop übertragen wird. Eingezeichnet ist nur eines der vielen Axone, die die Signale von der Retina zum Schwerpunktmodul übertragen. Dies dient der Übersichtlichkeit.
Genaugenommen liegen zwei solcher Schichten in der Retina vor, denn es gibt die Hell-On-Ganglienzellen und die Dunkel-On-Ganglienzellen. Dies ist in der obigen Abbildung vereinfachend weggelassen worden.
Ebenso weggelassen sind die Schichten mit den farbsensitiven Rezeptoren.
Insofern besteht ein komplettes Schwerpunktmodul im visuellen Bereich aus den gleichen Schichten wie der visuelle Thalamus. Wir betrachten hier beispielhaft nur eine der Schichten, um die prinzipielle Funktionsweise des visuellen Schwerpunktmoduls zu erläutern.
Wenn ein visuelles Objekt eine dieser Zellen besonders stark erregt, weil es von ihrem großen rezeptiven Feld wahrgenommen wird, gelangt diese Erregung über Axone des Sehnervs in das sensorische Zentrum des dritten Segments. Dort existiert eine retinotop geordnete Abbildung der Retina. Das Inputneuron der Klasse 4, welches den starken Input aus der Retina erhält, erregt die Interneuronen in der Schicht 4. Die zugeführte Erregung breitet sich in der Fläche aus, wobei eine abstandsabhängige Dämpfung auftritt.
Eine laterale Hemmung der Retinaneuronen untereinander verstärkt die Wirkung dieser Schaltung.
Wie denken uns sechs Outputneuronen in Form eines Sechsecks am äußeren Rand des sensorischen Kerns, der hier eine Fläche bildet. Diese sechs Outputneuronen versorgen mit ihren Signalen die sechs Augenmuskeln des Auges dieser Körperseite. Im Prinzip existiert diese Schwerpunktschaltung im Tectum opticum, beim Menschen als Colliculi superiores bezeichnet.
Befindet sich das Abbild des visuellen Objekts in der Mitte der Retina, so wird die dortige Ganglienzelle zur Mitte des Schwerpunktmoduls projizieren. Dann werden alle sechs Steuerneuronen für die Augenmuskeln gleich stark erregt, denn ihr Abstand zur Mitte ist gleich groß.
Befindet sich das Abbild des Objekts außerhalb der Mitte, so wird ein dort befindliches Retinaneuron maximal erregt und zieht zur zugeordneten Stelle des Schwerpunktmoduls. Diese Stelle liegt nun auch außerhalb der Mitte an der gleichen Position wie in der Retina.
Das Steuerneuron, welches den kleinsten Abstand zu diesem Schwerpunktneuron besitzt, erhält den stärksten Erregungsanteil, denn die Dämpfung auf der kurzen Strecke ist kleiner. Alle übrigen 5 Steuerneuronen des Schwerpunktmoduls erhalten eine abgeschwächte Erregung, die mit wachsendem Abstand zum Schwerpunktneuron abnimmt.
Daher erhalten die Augenmuskeln unterschiedlich starke Kontraktionssignale. Am stärksten kontrahiert der Augenmuskel, der die kleinste Entfernung zum Schwerpunktneuron besitzt. Dadurch wird das Auge so bewegt, dass das visuelle Objekt sich zur Mitte bewegt.
Hierbei sollten wir uns die Muskeln ähnlich vorstellen wie kleine Schrittmotoren. Jedes Aktionspotential führt zu einer kleinen Muskelverkürzung. Daher wird der Augenmuskel, welcher die höchste Feuerrate empfängt, die größte Verkürzung aufweisen. Die Augenbewegung führt dazu, dass das Abbild des Objekts sich der Netzhautmitte nähert.
Erst wenn alle Augenmuskeln die gleichen Kontraktionssignale empfangen, weil das Netzhautabbild des Objekts sich in der Netzhautmitte befindet, also zentriert dargestellt ist, erhalten alle Muskeln gleichstarke Kontraktionssignale, so dass keine Augenbewegung stattfindet, sondern nur noch die Muskelspannung aufrecht erhalten wird.
In der Abbildung 36 ist nur eine visuelle Modalität dargestellt. In der Realität sind hier die verschiedenen visuellen Modalitäten in Schichten übereinander angeordnet, etwa so wie im visuellen Thalamus. Dort findet man (beim Menschen) die magnocellularen Modalitäten Hell-On und Dunkel-On sowie die parvocellularen Modalitäten Rot-On/Grün-Off, Grün-On/Rot-Off sowie die Modalität Blau-On/Gelb-Off. Daher kann eine Augenbewegung zum Fokussieren sowohl von hellen, von dunklen und von farbigen Objekten ausgelöst werden. Hier setzt sich das stärkste Signal durch.
Wenn anstelle der Augenmuskeln diejenigen Halsmuskeln angesteuert werden, die eine Kopfdrehung bewirken, so kann die Augendrehung durch eine Kopfdrehung ergänzt werden. Auch die Ansteuerung von Rumpfmuskeln, die eine Körperdrehung bewirken, ist möglich. Dazu bilden die Eingangsneuronen für diese Modalitäten Schichten für die verschiedenen Muskelbereiche bzw. die zugehörigen Segmente.
Die sensorischen Module zur Augendrehung, Kopfdrehung und Rumpfdrehung sind einfach übereinander angeordnet werden, wobei sich die Retinasignale dann von oben nach unten durch diese Modulschichten durcharbeiten, hierzu verlaufen die Axone senkrecht durch die verschiedenen Schichten. Hierbei würden die Signale für die Steuerung der Augenmuskeln zuerst generiert werden, dann – mit geringer Verzögerung – die Halsmuskeln und zuletzt die Rumpfmuskeln. Das Ganze wäre ein dynamischer Prozess, weil bereits die ersten Outputsignale zu Augenbewegungen führen, die ihrerseits eine Neupositionierung des visuellen Objektabbildes bewirken.
Wenn die Signale des Retinaabbildes zusätzlich das Basalganglienmodul durchlaufen und als hemmende und zeitverzögerte Signale an den ursprünglichen Signalneuronen des Moduls andocken, können die Augen (unter Einbeziehung von Hals- und Rumpfmuskeln) auch bewegten Objekten folgen. Denn stillstehende Objekte löschen sich dann koplett aus, weil deren Vergangenheitsabbild mit dem Gegenwartsabbild identisch ist.
Möglich wurden Schwerpunktmodule dieser Art dadurch, dass der ursprünglich recht große Hohlraum des Neuralrohrs im Kopfbereich, der mit Ventrikelflüssigkeit gefüllt war, seinen Durchmesser verkleinern konnte, da es durch die Aufspaltung des Neuralrohrs auf der Höhe des Cerebellums nun auch einen äußeren Ventrikelraum gab, der die Stoffwechselprodukte der Nervenzellen fortleiten konnte. So verschwand der Ventrikelraum über dem Thalamus fast vollständig, so dass diese Art von Schwerpunktmodulen entstehen konnte. Bei einfachen Wirbeltieren bildet das Tectum opticum noch einen Abschnitt des hohlen Neuralrohrs, dessen Inneres vom Ventrikelraum gebildet wird.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan