Gehirntheorie des Menschen
ISBN 978-3-00-068559-0
Monografie von Dr. rer. nat. Andreas Heinrich Malczan
8. Das Speichermodul im Pontocerebellum
Das Cerebellum entstand sehr früh in der Evolutionsgeschichte und steht für den Beginn des Übergangs der Bilateria zu den Chordaten. Das Spinocerebellum steht sogar für den Übergang von den wasserbewohnenden Tieren zu den Landtieren, ermöglichte es doch durch die Einführung der Co-Aktivierung der motorischen Gegenspieler eine Stabilität des Körpers, die auch die Schwerkrafteinwirkung berücksichtigte.
Die Entstehung des Cerebellums wurde in meiner Monografie -Gehirntheorie der Wirbeltiere- ausführlich dargestellt. Natürlich ist meine Darstellung dort als Hypothese aufzufassen, für die es jedoch sehr viele Indizien gibt.
Wir müssen, um die Entstehung des Pontocerebellums verstehen zu können, zurück zu den Anfängen der Entwicklung des zentralen Nervensystems der einfachen, segmentierten Bilateria. Dort gab es in jedem Segment pro Körperseite je ein sensorisches und ein motorisches Zentrum. Mittelwertneuronen der Klasse 6 sammelten die vorhandene Erregung mit ihren größeren Dendritenbäumen ein und gewannen so Mittelwertsignale für die verschiedenen Mittelwertzentren.
Mit der Herausbildung der Nachbarhemmung konkurrierten auch die Mittelwertsysteme der verschiedenen Segmente miteinander, viele fielen dem neuronalen Konkurrenzkampf zum Opfer. Übrig blieb beispielsweise in der siebenten Etage des segmentierten Nervensystems das dopaminerge Mittelwertsystem der Substantia nigra pars compacta mit der Unterabteilung der VTA sowie die Formatio reticularis, die sich wahrscheinlich über mehrere Segmente erstreckt.
Im zweiten Segment, direkt unterhalb des Thalamus, konnte sich ein Mittelwertkern behaupten, der sich zum Nucleus subthalamicus entwickelte. Er empfing den Output der Mittelwertneuronen der Klasse 6 aus dem sensorischen und dem motorischen Kern der ersten Etage, die zum Cortex wurde.
Sein Output diente der Steuerung der verschiedenen Lebenserhaltungssysteme. Dazu mussten seine Signale abwärts steigen und erreichten so (unter anderem) den Ausgangskern des Urhirns in der siebenten Etage. Dieser Kern war der Nucleus ruber.
Der Nucleus ruber projizierte jedoch nicht nur absteigend zu den Motoneuronen des Rumpfes, sondern auch recht früh zum Seitenwechselkern dieser Etage (dieses Segments). Dieser Kern war der Nucleus olivaris. Zweck der Projektion war zunächst die kontralaterale Hemmung, denn anfangs standen beide Körperhälften in neuronaler Konkurrenz zueinander. Diese wurde durch hemmende Interneuronen des kontralateralen Nucleus ruber bewirkt, die von der Gegenseite erregt wurden.
Später, als sich diese hemmenden Interneuronen zu einem neuen Kern, dem Nucleus Purkinje separiert hatten, bewirkte dieser die kontralaterale Hemmung.
Seine Signale erreichten jedoch irgendwann die Formation reticularis. Diese war ein Mittelwertkern in dieser Etage. Die Hemmung der Mittelwertsignale durch den Purkinjekern führte zur Signalinversion genau dieser Signale. Die Neuronen der Formatio reticularis, die den hemmenden Input vom Purkinjekern empfingen, ihn invertierten und zum Nucleus ruber schickten, bildeten recht bald einen eigenen Kern. So entstand der erste Kleinhirnkern. Von da an kann man den Purkinjekern als Cerebellum bezeichnen, der Kleinhirnkern wird als Subsystem dieses Cerebellums angesehen.
Das Cerebellum versorgte nunmehr die Gegenseite des Körpers mit invertierten, aber erregenden Signalen. Die kontralaterale Hemmung der motorischen Gegenspieler wurde abgelöst durch die Co-Aktivierung, also die inverse Erregung. So waren beide Muskeln eines einfachen Gelenks angespannt, der eine mehr, der andere weniger. Dies galt zunächt nur für die Rumpfmuskulatur, bei der sich die motorischen Gegenspieler (meist) auf der anderen Körperseite befanden, die wurm- oder fischähnlichen Körper bewegten sich durch Schlängelbewegungen durch das Wasser der Urozeane. Durch die inverse Erregung der motorischen Gegenspieler waren beide Muskeln des Gelenks angespannt, der eine mehr, der andere evtl. weniger. Dadurch konnten beliebige Gelenkwinkel eingestellt werden auch unter äußerer Belastung, etwa durch die Schwerkraft. Erst da konnten die (künftigen) Tetrapoden das Wasser verlassen und an Land gehen.
Bei den Tetrapoden garantierte das
Spinocerebellum die gegenläufige motorische Anteuerung der
Extremitätenmuskeln, was die Fortbewegung vereinfachte.
Das Cerebellum empfing von Nucleus ruber seinen kompletten Signalsatz - das war die einfachste Lösung. Da ein Teil der Signale aus den darüberliegenden Mittelwertkernen stammte, kamen auch die Mittelwertsignale des Nucleus subthalamicus zum Nucleus ruber und von dort über den Olivenkern zum Cerebellum.
Nun entstand im Verlauf der Evolution bei Signalen und Neuronen die Tendenz, sich nach Modalitäten zu ordnen. Hier wurden die Neuronen der verschiedenen Schichten ab einer gewissen Entwicklungsstufe auch als verschiedene Modalitäten eingestuft und nahmen am Prozess der Trennung der Modalitäten teil.
So kam es, dass sich die Signale in der Olive, dem Nucleus olivaris, nach der Art ihrer Modalitäten anordneten. Die Segmentaufteilung des Körpers wurde beibehalten. Dies verdeutlicht die folgende Abbildung.
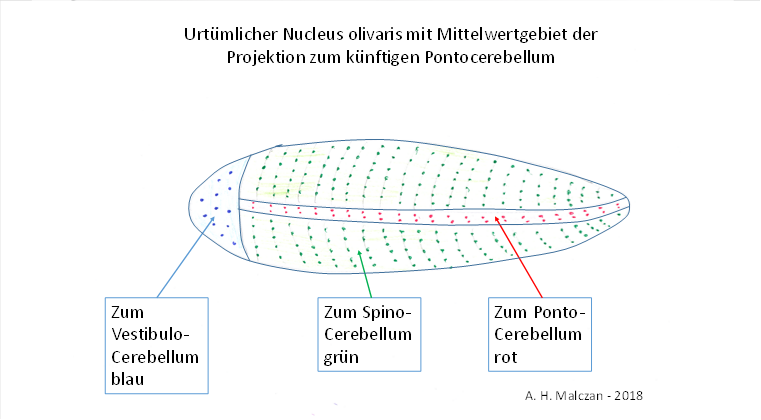
Abbildung 37: Urtümlicher Nucleus olivaris mit dem Mittelwertgebiet der Projektion zum künftigen Pontocerebellum
Im urtümlichen Nucleus olivaris begannen die Neuronen der Signalklasse 6, welche den Mittelwertsignalen zugeordnet sind, sich in zwei Streifen anzuordnen. Diese Mittelwertneuronen waren stark in der Unterzahl. Die Mittelwertsignale der Klasse 6 entstanden in jedem neuronalen Zentrum eines jeden Segments. Auch sie konnten im Verlauf der Evolution die Segmentgrenzen überwinden. So konnten sie sowohl aufsteigend als auch absteigend alle anderen Segmente erreichen. Aufsteigend erreichten sie auch das oberste Segment, welches der spätere Cortex wurde, zogen zur motorischen Seite und stiegen wieder herab. So verfügte auch der Cortex über alle diese Mittelwertsignale.
Absteigend erreichten diese Signale den Nucleus ruber, der sie jedoch auch der Olive übergab. Dort endeten diese Mittelwertsignale an Umschaltneuronen, die einen waagerecht verlaufenden und recht schmalen Neuronenstreifen bildeten, wie er in obiger Abbildung dargestellt ist.
Der Nucleus olivaris war also ein Körpermodell der signalliefernden Rezeptoren, er war wie der Körper segmentiert. Er war ebenfalls bilateral symmetrisch, je eine Hälfte repräsentierte eine Körperhälfte. Als der Körper der Tiere noch wurmförmig war, sah er ebenfalls wurmförmig aus. Und pro Körperhälfte empfing er in jedem Segment auch die Mittelwertsignale, die sich nun in einem kleinen Raumgebiet zusammenfanden, da sie in der Minderheit waren. So entstand ein kleiner Streifen von Neuronen in Nucleus olivaris, der die Mittelwertsignale der verschiedenen Segmente empfing.
Diese Neuronen projizierten ins Cerebellum und bildeten dort links und rechts (also für jede Körperhälfte) ebenfalls einen solchen Streifen. Damals ähnelten sich der Nucleus olivaris und das Cerebellum noch sehr, beide waren ebenso wurmförmig wie der Körper der damaligen Tiere. Aus dem schlanken Streifen der Mittelwertsignale ging das Pontocerebellum hervor. Dies ist meine feste Überzeugung. Viele Gründe sprechen dafür.
Im Zuge der Erweiterungsphasen des Cerebellums entstand zusätzlich zur Kletterfaserprojektion von der Olive eine Moosfaserprojektion. Der Input der Moosfasern kam aus dem Cortex über die Brückenkerne, wo sie die Seite wechselten. Damit entstammte der Moosfaserinput der Cortexseite, die auch die Kletterfasern speiste. Es muss als großer Fortschritt gelten, dass das Cerebellum nun auch die absteigenden Signale der Cortexrinde empfangen konnte.
Hier wurde die Topologie des Cortex in das Pontocerebellum übernommen.
Im Cortex gab es das parvocellulare Teilsystem, welches auf der sensorischen Seite aus den Schichten 3 und 4 gebildet wurde. Auf der motorischen Seite bestand das parvocellulare System aus den Schichten 2 und 5. Die Schicht 1 des Cortex war eine reine Inputschicht.
Das magnocellulare System bestand aus der Mittelwertschicht 6, die sowohl sensorisch als auch motorisch vertreten war.
Wir analysieren die cortikale Projektionen zum Pontocerebellum. Das parvocellulare System projiziert über die Brückenkerne in das Pontocerebellum, wo die Signale über das Moosfasersystem die Körnerzellen versorgen, deren Axone die Purkinjezellen rechtwinklig kreuzten und direkt oder über Interneuronen kontaktieren.
Die Schicht 6 des sensorischen und motorischen Cortex projiziert zum Mittelwertkern der thalamischen Etage, also zum Nucleus subthalamicus. Dort integrieren große Mittelwertneuronen die Erregung, wobei jeweils mehrere benachbarte Mittelwertneuronen des Cortex in ein gemeinsames Mittelwertneuron des Nucleus subthalamicus projizieren. Das Cortexgebiet, welches über die cortikalen Mittelwertneuronen in ein gemeinsames Mittelwertneuron des Nucleus subthalamicus projiziert, werden wir in dieser Monografie als Cortexcluster bezeichnen. Ihm ist genau ein Mittelwertneuron im Nucleus subthalamicus zugeordnet. Dessen Mittelwertsignal zweiter Stufe erreicht das Cerebellum über den Umweg vom Nucleus ruber zur Olive und von dort als Kletterfaser zu den Purkinjezellen.
Jeder Purkinjezelle ist eine Kletterfaser zugeordnet, die den Signalmittelwert eines Cortexclusters repräsentiert. Die Cortexneuronen der Klasse 3 projizieren über die Brückenkerne und die Moosfasern in ein Cerebellumgebiet, in dem sich - unter anderem - genau diese Purkinjezelle befindet. Wir werden dieses Gebiet als Cerebellumcluster bezeichnen.
Weiterhin einigen wir uns darauf, die Signale der cortikalen Neuronen der Klasse 3 als Elementarsignale zu bezeichnen.
Dann projizieren die cortikalen Elementarsignale in die Körnerzellen eines Cerebellumclusters, während ihr Mittelwertsignal zweiter Stufe die Kletterfasern des Cerebellumclusters speist. Das Mittelwertsignal erster Stufe entsteht in der sechsten Schicht des Cortex durch die dortigen magnocellularen Pyramidenzellen. Die zweite Mittelwertstufe erzeugt der Nucleus subthalamicus.
Die cortikalen Mittelwertsignale gewannen im Verlauf der Evolution an Stärke, weil die rezeptiven Felder größer wurden und die Anzahl der Cortexneuronen (bei vielen Spezies) fast explosionsartig anstieg. Dies trifft vor allem bei den Primaten zu, ganz besonders jedoch beim Menschen. Dennoch begann das Pontocerebellum schon viel früher seine Arbeit im Dienst der Wirbeltiere. Bei vielen einfachen Tieren ist der Cortex noch recht klein, doch ihr Cerebellum bildet bereits eine prominente Struktur beachtlicher Größe.
Da der Nucleus subthalamicus eine weitere Mittelwertbildung aus dem Output der Neuronenklasse 6 vornahm, stiegt die mittlere Feuerrate seiner Outputneuronen im Verlauf der Evolution immer stärker an. So empfing jede Purkinjezelle - falls ihr Cortexcluster über aktive Neuronen verfügte - eine starke Mittelwerterregung. Und in vielen Kopfsegmenten waren fast immer Neuronen aktiv, da es ständig Umweltsignale olfaktorischer, visueller, vestibulärer oder taktiler Art gab.
Nun führt ein langanhaltendes, stärkeres Signal zur Überlastung der Neuronen. Daher war es günstig, wenn Signalpausen eingeschoben wurden. Diese Signalpausen entstanden durch eine hemmende Projektion aus dem Striatum.
Wir gehen davon aus, dass die Mittelwertneuronen des Nucleus subthalamicus auch in den dopaminergen Mittelwertkern der siebenten Etage projizierten, denn jedes Signal sollte bei Mittelwertkernen berücksichtigt werden. Daher empfing die Substantia nigra pars compacta diese Mittelwertsignale zweiter Stufe. Sie behandelte diese Signale so wie alle Signale. Sie wurden auf Dopamin umgeschaltet und zum Striatum gesendet, wo sie an hemmenden Striosomenneuronen endeten und diese erregten. Diese Striosomenneuronen waren nun erregt, solange die Substantia nigra sie erregte.
Nun entwickelte sich im Verlauf der Evolution auch eine hemmende Rückprojektion vom Striatum zur Substantia nigra pars compacta. Natürlich brauchten die hemmenden Signale eine gewisse Zeit, bis sie vom Striatum aus die Substantia nigra erreichten. In dieser Zeit konnten die Striosomensignale die Mittelwertneuronen des Nucleus ruber hemmen, so dass die zugehörigen Kletterfasersignale zum Stillstand kamen, weil sie gehemmt wurden.
Wenn jedoch die hemmenden Striosomensignale die Substantia nigra pars compacta endlich erreichten, konnten sie diese völlig hemmen. Ab diesem Zeitpunkt konnte die Substantia nigra keine Aktionspotentiale zu den Striosomen senden. Waren diejenigen, die bereits unterwegs waren, aufgebraucht, so empfingen die Striosomen keine erregenden Signale mehr, ihre Neuronen verstummten. Dadurch fiel die Hemmung des Nucleus ruber weg, so dass die Mittelwertsignale des Nucleus subthalamicus vom Nucleus ruber zum Nucleus olivaris und somit zu den Purkinjezellen durchgelassen wurden.
Da jedoch gleichzeitig die Substantia nigra vom Nucleus subthalamicus erregt wurde, produzierte sie wieder erregende Aktionspotentiale, die nach einer gewissen Laufzeit in den Striosomen ankamen und dort hemmende Aktionspotentiale in Richtung der Substantia nigra und in Richtung des Nucleus ruber sandten. So wurde die Signaldurchlässigkeit des Nucleus ruber mittels der Striosomen ständig kurzzeitig unterbrochen und danach wiederhergestellt. So entstand die typische Schwingungsform der Kletterfasersignale im Pontocerebellum: Eine tetanische Schwingung, die laufend von kurzen Pausen unterbrochen wird. So wurde eine Überlastung der beteiligten Neuronen vermieden.
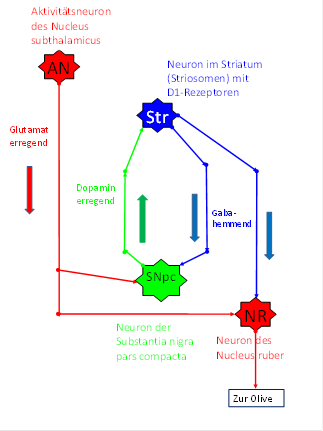
Abbildung 38: Periodische Hemmung der Mittelwert-Kletterfasersignale durch das Striatum
Damit erhielten die Kletterfasersignale im Pontocerebellum folgendes Aussehen:

Abbildung 39: Kletterfasersignale im Pontocerebellum - prinzipielles Aussehen
Ein starkes Mittelwertsignal - immer dort vorhanden, wo starke cortikale Aktivität auftrat - konnte in einer Purkinjezellen den Prozess der Langzeitpotenzierung (LTP) und der Langzeitdepression (LTD) verursachen. Dieser Prozess fand statt, wenn die Purkinjezelle gleichzeitig zweierlei Input empfing:
· Cortikalen Input über die Parallelfasern.
· Tetanischen (höherfrequenten) Input über die Kletterfasern.
Denn ein tetanischer Kletterfaserinput bewirkte in recht kurzer Zeit (im Sekundenbereich) die Erhöhung bzw. die Verminderung der synaptischen Kopplung genau bei denjenigen Synapsen, die gleichzeitig vom cortikalen Input erregt wurden. Ein Umbauprozess in den Synapsen bewirkte eine bleibende Veränderung der Kopplungsstärke.
Die Veränderung der Kopplungsstärke blieb dauerhaft erhalten. Dies lag an Umbauarbeiten in der Zelle selbst, deren Start durch die tetanische Erregung angestoßen wurde. Die bleibende Veränderung der synaptischen Stärke wollen wir in dieser Monografie als Prägung bezeichnen.
Trat nach der Prägung genau dasjenige Signal auf, welches während des Prozesses der LTP oder LTD anlag, so reagierte die Purkinjezelle mit einer deutlichen Veränderung ihrer Feuerrate. Bei LTP feuerte sie nun viel stärker, bei LTP deutlich schwächer.
Das Cerebellum wurde also lernfähig.
Doch wie konnte gesteuert werden, welche Parallelfasern an der LTP teilnahmen und welche an der LTD?
Hierzu müssen wir den Aufbau von Cortex und Pontocerebellum aus Clustern berücksichtigen.
Für die Analyse der Topologie der Cortex- und Cerebellumcluster gehen wir davon aus, dass sich die Axone der Körnerzellen bereits T-förmig verzweigten und sich parallel zueinander ausrichteten, so dass sie die Ebene der Dendritenbäume der Purkinjezellen im rechten Winkel kreuzten.
Für die Analyse der Signalprojektion vom Cortex zum Pontocerebellum nutzen wir das Clustermodell. Ein Cluster (sowohl im Cortex als auch im Cerebellum) sei vereinfacht ein quadratischer Ausschnitt der Rinde. Ein Cortexcluster enthält eine größere Anzahl von Mittelwertneuronen der Klasse 6, die in der sechsten Schicht des Cortex mit ihren großen rezeptiven Feldern die Signale der Klassen 5 oder 3 aufsaugen und daraus eine Mittelwertaktivität erzeugen. In sensorischen Cortexfeldern sind es die Neuronen der Klasse 3, die die Mittelwertneuronen der Klasse 6 speisen. In motorischen Cortexfeldern - und jeder Gyrus hat einen sensorischen und einen motorischen Anteil - sind es die Neuronen der Klasse 5, die die Mittelwertneuronen der Klasse 6 aktivieren.
Eine größere Anzahl der Mittelwertneuronen aus einem ebenfalls größeren Cortexgebiet sendet nun die Axone der Mittelwertneuronen der Klasse 6 zu einem gemeinsamen Mittelwertneuron des Nucleus subthalamicus. Dieses spezielle Neuron ist unser Mittelwertneuron der Stufe 2.
Nun können wir genau definieren, was ein Cortexcluster ist:
Ein Cortexcluster ist ein Cortexgebiet, dessen Mittelwertneuronen der Klasse 6 auf genau ein Neuron des Nucleus subthalamicus konvergieren.
Beispielhaft könnte im primären visuellen Thalamus ein Cortexcluster aus 25 Helligkeitsmodulen mit räumlicher Signalausbreitung bestehen. Dann wäre ein Quadrat aus 5 benachbarten Modulen in der Breite und 5 benachbarten Modulen in der Länge ein Cortexcluster.
Wir setzen hier (hypothetisch) voraus, dass die Outputsignale der verschiedenen visuellen Module ebenfalls wie verschiedene Modalitäten behandelt werden und ihre Mittelwertneuronen sich im Nucleus subthalamicus ebenfalls nach Modalitäten aufspalten (Hell/Dunkel, Farbe, Linienelemente, -).
Diese verschiedenen Modalitäten besitzen sowohl im Nucleus subthalamicus als auch im Pontocerebellum eigene, zusammenhängende Bereiche.
Wir betrachten nun solche cortikalen Cluster und ihre Eigenschaften.
Benachbarte Cluster seien untereinander etwa gleichgroß. Wir können benachbarte Cluster in Clustergruppen zusammenfassen.
Eine Clustergruppe bestehe beispielhaft aus 9 Clustern und sei ebenfalls quadratisch. Der Cluster mit der Nummer 5 sei der Innencluster, die übrigen nennen wir Außencluster. Hierbei seien die Cluster 1, 3, 7 und 9 die Eckcluster. Die Richtung, in der die Cluster 4, 5 und 6 angeordnet sind, bezeichnen wir als horizontal, sie sind in einer Reihe angeordnet.
Die Richtung der Cluster 2,5 und 8 nennen wir vertikal, diese Cluster bilden eine Spalte.
In der folgenden Abbildung ist eine Clustergruppe dargestellt, die 9 Cluster sind wie beschrieben nummeriert.
In der Clustergruppe des Cerebellums mögen die Parallelfasern von oben nach unten verlaufen, wenn die untere Abbildung die Cortex- und Cerebellumrinde von oben zeigt.
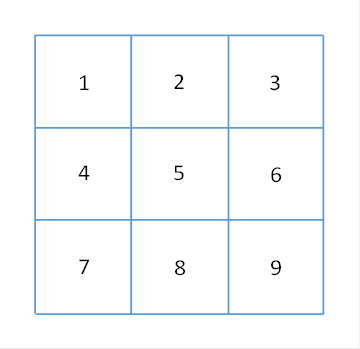
Abbildung 40: Clustergruppe im Cortex
Wir bezeichnen den Cluster 5 als Innencluster und die übrigen als Außencluster oder Randcluster der Clustergruppe. Jedem Cluster können wir ein Signal zuordnen, so gehört das Signal S5 zum Cluster 5.
Innerhalb eines Clusters seien die Signalneuronen annähernd gleichabständig verteilt. Diese Signalneuronen gehören auf der sensorischen Cortexseite zur Neuronenklasse 3, auf der motorischen zur Klasse 5. Ihre Signale bezeichnen wir als cortikale Elementarsignale. Sie projizieren über die Brückenkerne zum Pontocerebellum.
Beispielhaft ist nachfolgend ein Cluster mit 25 Outputneuronen dargestellt, die in 5 Reihen zu je 5 Neuronen angeordnet sind. Die reale Anzahl der Signalneuronen im Cortex ist bei höheren Wirbeltieren sicherlich um ein Vielfaches höher. Hier geht es nur um das Organisationsprinzip der
Clustertopologie.

Abbildung 41: Aufbau eines Clusters aus Neuronen im Cortex
Es wird deutlich, dass ein Clustersignal, etwa S5, sich aus vielen Elementarsignalen zusammensetzt, die man in Spalten und Zeilen anordnen und durchindizieren kann. Gehört zum Cluster 5 in der zweiten Zeile und vierten Spalte der Output O5,2,4, und stellt die Zahl 5 die Anzahl der Zeilen und der Spalten dar, so kann (stark vereinfacht) das Signal S5 als Matrix geschrieben werden:
|
s5,1,1 |
s5,1,2 |
s5,1,3 |
s5,1,4 |
s5,1,5 |
|
s5,2,1 |
s5,2,2 |
s5,2,3 |
s5,2,4 |
s5,2,5 |
|
s5,3,1 |
s5,3,2 |
s5,3,3 |
s5,3,4 |
s5,3,5 |
|
s5,4,1 |
s5,4,2 |
s5,4,3 |
s5,4,4 |
s5,4,5 |
|
s5,5,1 |
s5,5,2 |
s5,5,3 |
s5,5,4 |
s5,5,5 |
= S5
Hierbei repräsentiert s5,k,l das Elementarsignal in der k-ten Zeile und der l-ten Spalte, welches im Cluster Nummer 5 vom Signalneuron in der k-ten Zeile und der l-ten Spalte geliefert wird.
Wir postulieren eine Signalprojektion der Signalneuronen der Cortexcluster über die Moosfasern in die Körnerzellen der Cerebellumcluster und gehen davon aus, dass diese Signalabbildung topologietreu sei in dem Sinne, dass die Clusternummer dabei erhalten bleibt.
Eine Besonderheit des Pontocerebellums muss am Anfang vorangestellt werden. Der Output des Pontocerebellums ist normalerweise gleich null. Denn die Erregung eines Neurons des Nucleus dentatus durch das über die Kletterfaser eintreffende Signal wird aufgehoben dadurch, dass auch die Purkinjezelle durch genau dasselbe Signal erregt wird. Beide besitzen eine gleichstarke Erregung. Doch die Purkinjezelle verwendet einen hemmenden Transmitter, daher hemmt sie das Outputneuron im Nucleus dentatus. Deshalb gibt es im Pontocerebellum normalerweise keinen Output.
Eine Struktur ohne Nutzen wäre auf die Dauer sicherlich eliminiert worden. Also muss es einen Nutzen geben. Dieser liegt in dem Umstand begründet, dass die Parallelfasern die Purkinjezellen direkt erregen können, aber ebenso auch indirekt über zwischengeschaltete Stern- und Korbzellen hemmen können.
Hier kommt der wichtige Aspekt der Signalverwandtschaft ins Spiel.
· Die Cortexneuronen der Schichten 3 bzw. 5 innerhalb eines Clusters sind signalverwandt. Diese Signalverwandtschaft entsteht über zwei Stufen.
· Diese Cortexneuronen versorgen jeweils (meist nur) ein Mittelwertneuron in dessen rezeptiven Feld.
· Dieses Mittelwertneuron versorgt mit allen anderen Mittelwertneuronen des Clusters genau ein Neuron im Nucleus subthalamicus.
· Signalverwandtschaft wird in Neuronenketten in beide Richtungen weitergegeben.
· Daher sind die Cortexneuronen eines Cortexclusters untereinander signalverwandt, weil es einen gemeinsamen Signalempfänger im Nucleus subthalamicus gibt.
In Neuronenketten wird Signalverwandtschaft in beide Richtungen weitergegeben.
Sind Neuronen nicht signalverwandt, so stehen sie in neuronaler Konkurrenz zueinander. Dann hemmen sie sich gegenseitig.
Daher können Parallelfasern, die Input aus den Randclustern empfangen, die Purkinjezelle im Innencluster hemmen, denn sie stehen in neuronaler Konkurrenz zueinander. Die Hemmung erfolgt durch zwischengeschaltete Interneuronen, die wir als Sternzellen und Korbzellen bezeichnen.
Und Parallelfasern, die Input vom Innencluster empfangen, erregen die Purkinjezellen des Innenclusters.
Dadurch kann das fein austarierte Gleichgewicht zwischen dem erregten Dentatusneuron und der zugehörigen hemmenden Purkinjezelle aus dem Gleichgewicht gebracht werden, woraufhin es plötzlich doch einen Output aus dem Pontocerebellum gibt.
Standardmäßig wird die Purkinjezelle jedoch von den Cortexsignalen des eigenen Clusters erregt, so dass sie das Outputneuron aus dem Nucleus dentatus trotzdem hemmt. Da jedoch das Dentatusneuron ebenfalls den Input aus dem Innencluster empfängt, heben sich beide Erregungsanteile auf, der Output ist das Nullsignal.
Nur, wenn gleichzeitig die Signale der Außencluster aus dem Cortex über die Parallelfasern eintreffen, hemmen sie die Purkinjezelle, weil sie die zwischengeschalteten Stern- und Korbzellen erregen. Dann bleibt im Dentatusneuron der Erregungsanteil übrig, der vom Innencluster herrührt. Es gibt einen erregenden Output zum Cortex.
Dies ist aus mathematischer Sicht interessant: Nur wenn Innensignale und Außensignale gleichzeitig einwirken, wird ein Output erzeugt.
Innensignal + Außensignal = Outputsignal.
Das Innensignal generiert keinen Output, das Außensignale allein kann dies ebenfalls nicht. Nur beide zusammen können dies.
Dies ist eine Signalverknüpfung über eine Und-Beziehung. Mathematiker bezeichnen dies als Konjunktion. So entstehen Komplexsignale. Komplexsignale setzen sich also immer zusammen aus einem Erregungsanteil vom Innencluster und einem Erregungsanteil von den Außenclustern.
Doch das Pontocerebellum konnte noch mehr: Es konnte diese Komplexsignale auch erlernen.
Erlernen bedeutet, dass das Cerebellum, welches ein Komplexsignal noch nicht kennt, darauf nur schwach oder gar nicht reagiert. Nachdem das Cerebellum dieses Komplexsignal erlernt hat, feuert es deutlich stärker, sobald dieses Komplexsignal als Input präsentiert wird.
Wir wollen den Prozess das Erlernen eines Signals im Pontocerebellum als Prägung bezeichnen. Diese Prägung erfolgt durch LTP und LTD, wodurch die synaptische Kopplung zwischen den beteiligten Neuronen dauerhaft und bleibend verändert wird.
Grundlage für die Prägung sind folgende Organisationsprinzipien:
· Die Topologie der Cortexcluster und der Cerebellumcluster.
· Die Nachbarhemmung von Signalen, die nicht (hinreichend) miteinander verwandt sind.
· Die tetanische Mittelwerterregung des Mittelwertneurons. Sie tritt in einem Cortexcluster auf, wenn eine Teilmenge von Neuronen im Cluster hinreichend aktiv ist.
· Den Prozess der Prägung (des Erlernens) durch LTP und LTD mit Hilfe dieses tetanischen Mittelwertsignals.
· Den Prozess der Entprägung (des Vergessens) mit Hilfe eines niederfrequenten Kletterfasersignals
Damit ist das tetanische Mittelwertsignal der Speicherbefehl, vergleichbar mit dem Speicherbefehl in herkömmlichen Computern, während das niederfrequente Mittelwertsignal dem Löschbefehl entspricht.
Wie kommt Signalverwandtschaft zustande? Sie wird in Neuronen von Neuron zu Neuron übertragen. Mittelwertsignale sind Abkömmlinge der Clustersignale. Daher besteht eine Signalverwandtschaft zwischen ihnen.
Es scheint - so meine Hypothese - bei Neuronen zwei Arten von Grundregeln zu geben:
· Errege den, der noch nicht erregt ist (falls er erreichbar ist).
· Hemme die Konkurrenz.
So entwickelten sich beispielsweise die Mittelwertneuronen. Jedes Neuron, welches aktiv war, während das Mittelwertneuron inaktiv war, konnte an ihm andocken und es nun ebenfalls erregen. Vorausgesetzt, sein Axon war lang genug. So ist jedes Neuron mit seinem Mittelwertneuron signalverwandt.
Besitzt ein Neuron dagegen einen hemmenden Transmitter, so kann es nur dann synaptischen Kontakt mit einem anderen Neuron aufnehmen, wenn beide gleichzeitig aktiv sind. Daher hemmt ein Neuron auch jedes Mittelwertneuron, in dessen Signalmenge sein eigenes Signal bereits vorhanden ist. Deswegen hemmen dopaminerge Signale auch die Mittelwertneuronen, zu denen sie selbst beigetragen haben. Doch nun weiter im Text zum Pontocerebellum.
Wie wirken nun die vier Organisationsprinzipien bei der Prägung im Pontocerebellum mit?
Wir unterstellen eine komplexere Form von LTP und LTD im Pontocerebellum, die dazu führt, dass ein Outputneuron des Nucleus dentatus auf eine bestimmte Signalkonstellation deutlich stärker reagiert. Diese Signalkonstellation bezeichnen wir als Prägungssignal. Den Vorgang der LTD und LTP bezeichnen wir als Prägung.
Das Prägungssignal entstamme einer Clustergruppe des Cortex und möge aus zwei Teilsignalen bestehen:
- Dem Innensignal Si aus dem Innencluster einer Clustergruppe.
- Dem Außensignal SA aus den Außencluster dieser Clustergruppe.
Das Innensignal darf nicht das Nullsignal sein, es muss eine ausreichende Teilmenge der Cortexneuronen aus dem Innencluster aktiv sein. Es bildet die Grundlage für den Signalmittelwert.
Das zugehörige Mittelwertsignal endet an der Purkinjezelle eines Cerebellum-Innenclusters und am zugehörigen Dentatusneuron.
Das Innensignal Si endet an erregenden Parallelfasern der Purkinjezelle des Cerebellum-Innenclusters und am zugehörigen Dentatusneuron.
Das Außensignal SA endet an hemmenden Stern- und Korbzellen des gleichen Cerebellum-Innenclusters. Die Dentatusneuronen des Innenclusters können von den Signalen der Außencluster nicht erregt werden, da die Entfernung zwischen den Neuronen viel zu groß ist.
Wenn das Outputneuron des Nucleus dentatus stärker auf das Komplexsignal S = Si + SA reagieren soll, muss die Purkinjezelle von diesen Signalen weniger erregt werden.
Dazu wäre Folgendes erforderlich:
- Das Innensignal liegt an Parallelfasern an, deren synaptischen Kopplung durch das tetanische Kletterfasersignal (LTD) vermindert wird, beispielsweise vom Kopplungswert 1 auf den Kopplungswert ½. So wird die Purkinjezelle vom Innensignal weniger erregt.
- Das Außensignal liegt an den Stern- und Korbzellen, die im Sinne einer lateralen Hemmung die Purkinjezelle hemmen. Die synaptische Kopplungsstärke möge den Wert k besitzen. Durch die Einwirkung der tetanischen Erregung durch das Kletterfasersignal wird die synaptische Kopplung zwischen den Stern- bzw. Korbzellen zur Purkinjezelle verstärkt (LTP), z. B. von k auf 2k. So wird die Purkinjezelle vom Außensignal stärker gehemmt. Dadurch wird sie noch weniger erregt sein.
- Dadurch wird das zugehörige Outputneuron in Nucleus dentatus viel weniger gehemmt, so dass sein Output deutlich stärker ausfällt.
- Da das tetanische Kletterfasersignal auch am Outputneuron des Nucleus dentatus endet, wird (wahrscheinlich) auch die synaptische Kopplung zwischen den aktiven Moosfasern des Komplexsignals und den Dentatusneuronen stärker (LTP), z. B. von 1 auf 2. Damit wird das Outputsignal zusätzlich stärker.
- Die durch das tetanische Kletterfasersignal veränderten synaptischen Kopplungswerte bleiben dauerhaft auf dem neuen Niveau, so dass ein identisches Moosfasersignal wie dasjenige zur Zeit der Prägung nun viel stärkere Outputsignale im Nucleus dentatus erzeugt.
Damit überlagern sich mehrere Effekte und bewirken, dass das erregende Outputneuron im Nucleus dentatus viel stärker auf das Komplexsignal reagiert als vor der Prägung, denn dieses Outputneuron wird stärker erregt und weniger gehemmt.
Tritt das Prägungssignal später noch einmal auf, so ist die Antwort deutlich stärker. Wir bezeichnen das Prägungssignal nach der Prägung als Eigensignal der Purkinjezelle auf.
Eine erneute Prägung durch das Mittelwertsignal ist bei dieser Purkinjezelle nicht möglich. Die Moosfasersignale haben eine deutlich kürzere Laufzeit über die Brückenkerne als das Kletterfasersignal über den Nucleus subthalamicus, den Nucleus ruber und den Nucleus olivaris. Dadurch wird die Purkinjezelle vom Moosfaserinput aus den Außenclustern bereits gehemmt, bevor das Kletterfasersignal eintrifft. Damit ist es wirkungslos, denn gehemmte Neuronen stellen für erregende Signale eine Art Kurzschluss dar.
In der Natur beobachten wir einen Hang zur Bildung von Reserven. Im Cerebellum wird dies realisiert durch die Zusammenschaltung von Purkinjezellen zu Purkinjegruppen. Eine Purkinjegruppe besteht meist aus bis zu drei Purkinjezellen, die das gleiche Prägungssignal erlernen. Dadurch kommt eine zusätzliche Sicherheit in das System.
Erkennbar ist eine Purkinjegruppe dadurch, dass sie mit einer Golgizelle endet. Alle Purkinjezellen einer Purkinjegruppe projizieren auf ein gemeinsames, erregendes Outputneuron im Nucleus dentatus. Die Funktion der Golgizellen wird später beschrieben werden.
Fassen wir zusammen:
Durch die Prägung mittels LTD und LTP wird das Prägungssignal einer cortikalen Clustergruppe zum Eigensignal der Purkinjezellen einer Purkinjegruppe des zugehörigen Cerebellumclusters. Diese reagieren nach der Prägung auf das nochmalige Auftreten ihres Eigensignals mit deutlich verminderter Hemmung des erregten Outputneurons im Nucleus dentatus, so dass dessen Output auf das Eigensignal deutlich stärker ausfällt. Dadurch kann das Pontocerebellum in jedem Cluster genau ein Prägungssignal pro Purkinjegruppe erlernen und später wiedererkennen. Der Erkennungsoutput erreicht den Cortex.
Jede Purkinjegruppe kann also gemeinsam mit ihrem Outputneuron im Nucleus dentatus genau ein Prägungssignal erlernen und als Eigensignal abspeichern, indem die synaptischen Kopplungsstärken für dieses Komplexsignal dauerhaft und bleibend verändert werden. Insofern sind die Synapsen der beteiligten Interneuronen des Cerebellums die materiellen Orte, an denen die Signalspeicherung erfolgt.
Anfänglich gab es pro Outputneuron im Nucleus subthalamicus genau eine Purkinjegruppe. Damit zum zugehörigen Cortexcluster mehr als nur ein Komplexsignal erlernt werden konnte, entwickelten sich zwei verschiedene Möglichkeiten.
Die erste Möglichkeit bestand darin, dass eine Kletterfaser mehrerer Purkinjegruppen kontaktierte, indem sie ihre Länge vergrößerte. Entlang dieser nun deutlich längeren Kletterfaser konnten mehrere Purkinjegruppen kontaktiert werden, ebenso mehrere zugehörige Neuronen des Nucleus dentatus. Wir werden diese Lösung als sequentielle Divergenz bezeichnen.
Im Verlaufe der Evolution kontaktierte eine Kletterfaser mehrere Purkinjezellen durch sequentielle Divergenz, ihre Anzahl nahm in der Evolutionsreihe langsam zu.
Dadurch entstand ein neues Problem, welches gelöst werden musste. Wenn Purkinjegruppen von der gleichen Kletterfaser kontaktiert werden und die gleichen cortikalen Signale über die Körnerzellen empfangen, dann werden sie alle gleichzeitig das gleiche Prägungssignal erlernen. Dies wäre eine Verschwendung von Ressourcen. Hier kommen die Golgizellen ins Spiel.
Wie kann die Entstehung der Golgizellen erklärt werden? Wessen Abkömmlinge sind sie?
Auch zwischen den Signalneuronen der Klasse 6 im Cortex gab es bereits seit Urzeiten eine laterale Hemmung zur Kontrastverstärkung des Outputs. Die Mittelwertneuronen der Klasse 6 übertrugen diese Clusterkonkurrenz auf die Neuronen der Klasse 5 der Cluster. Diese neuronale Konkurrenz zwischen den Clustern wurde von den Moosfasern übernommen und an die Körnerzellen weitergereicht. Daher hemmten Körnerzellen, die zu einem Cerebellumcluster gehörten, die Körnerzellen der Nachbarcluster. Die dazu nötigen Interneuronen können als Abkömmlinge der hemmenden Interneuronen des Cortex interpretiert werden, die die Clusterhemmung realisierten.
Golgizellen sind Abkömmlinge der hemmenden Interneuronen der cortikalen Etage, Moosfaserneuronen sind die Abkömmlinge der cortikalen Signalneuronen. Körnerzellen sind Abkömmlinge der Moosfaserneuronen.
Die laterale Hemmung zwischen den cortikalen Signalen im Moosfasersystem erfolgt durch hemmende Interneuronen, deren Abkömmlinge im Körnerzellsystem die Golgizellen darstellen.
Der Körnerzellinput eines Clusters erregte die Golgizellen dieses Clusters, die ihrerseits mit ihren Axonen an den Dendriten der Körnerzellen der Nachbarcluster andockten und sie hemmten, falls sie selbst erregt waren. So wurde die gegenseitige Hemmung der Cluster im Pontocerebellum realisiert. Im Spinocerebellum und im Vestibulocerebellum gab es eine analoge Entwicklung. Doch im Pontocerebellum ermöglichte dieser Algorithmus die Verhinderung der sogenannten Mehrfachprägung. Diese wird im nachfolgenden Text erläutert.
Die Zahl der Golgizellen ist etwas kleiner als die der Purkinjezellen. Wir unterstellen hier beispielhaft, auf eine Golgizelle kämen drei Purkinjezellen.
Golgizellen werden vom Kletterfasersignal erregt und von Purkinjezellen gehemmt. Denn das Kletterfasersignal ist der Mittelwert der Clustersignale, und diese erregen ja die Golgizelle.
Vereinfacht gesagt: Golgizellen und Purkinjezellen sind signalverwandt, daher nehmen sie kontakt auf.
Wir betrachten beispielhaft eine Gruppe aus drei hintereinander angeordneten Purkinjezellen, die nacheinander von der gleichen Kletterfaser kontaktiert werden. Als viertes Neuron möge die Kletterfaser eine Golgizelle kontaktieren. Der weitere Verlauf der Kletterfaser möge zunächst nicht interessieren.
Wir unterstellen, dass die drei Purkinjezellen mit einem Prägungssignal S1 geprägt werden. Sie mögen wegen ihres identischen Outputs genau ein gemeinsames Outputneuron im Nucleus dentatus aktivieren. Somit gibt es für diese drei Purkinjezellen nur ein gemeinsames Outputneuron im Nucleus dentatus. Für jedes geprägte Signal gibt es also zwei Reserve-Purkinjezellen, falls eine ausfällt, bleibt die Schaltung funktionsfähig. Nur der Ausfall aller drei Purkinjezellen führt zum Verlust des erlernten Eigensignals.
Purkinjezellen der gleichen Purkinjegruppen enden jeweils mit einer Golgizelle und werden von der gleichen Kletterfaser aktiviert. Sie werden identisch geprägt. Ihr Output konvergiert auf das gleiche Dentatusneuron. Dieses meldet die Erkennung des geprägten Eigensignals an den Cortex.
Jede Golgizelle ist synaptisch mit der Kletterfaser verbunden, die die davor angeordneten Purkinjezellen kontaktiert. Während der Prägung aktiviert diese Kletterfaser mit ihrem starken Kletterfasersignal auch die Golgizelle, die stark erregt wird und den Signalfluss zu den Körnerzellen unterbricht. Damit erhalten die nachfolgenden Purkinjezellen weniger starke Parallelfasersignale, weil eine ganze Population von Parallelfasern (die Fremdsignale) quasi abgeklemmt wird. Ohne die Einwirkung von Fremdsignalen ist jedoch der Output einer Purkinjezelle gleich Null.
Nach einer Prägung erkennen Purkinjezellen während der Schwingungspause der Kletterfasersignale das anliegende Prägungssignal, da es ihr Eigensignal wurde. Der Hemmungswegfall führt zu einem starken Outputsignal des zugehörigen Neurons im Nucleus dentatus.
Hier kommt die hemmende Rückwirkung des Nucleus dentatus auf den Nucleus olivaris zur Geltung. Jedes erregende Outputneuron des Nucleus dentatus aktiviert mit seiner Erregung auch ein hemmendes Projektionsneuron, dessen Axon zum Nucleus olivaris zieht und dort genau an dem Inputneuron andockt, welches das prägende Kletterfasersignal erzeugt hat. Damit wird das Kletterfasersignal stark geschwächt, so dass nachfolgende Purkinjegruppen nicht mehr mit diesem Komplexsignal geprägt werden können.
Die hemmende Rückprojektion des Nucleus dentatus zum Nucleus olivaris dient der Verhinderung der Mehrfachprägung. Ohne Golgizellen und ohne diese hemmende Rückkopplung würden alle Purkinjegruppen eines Cerebellumclusters mit dem gleichen Prägungssignal geprägt werden.
Nun konnte das Pontocerebellum pro Cortexcluster mehrere, voneinander verschiedene Komplexsignale mittels LTD und LTP erlernen, weil es gelang, die Mehrfachprägung verschiedener Purkinjegruppen einer Kletterfaser mit einem identischen Prägungssignal zu verhindern.
Jede geprägte Purkinjegruppe reagiert mit deutlich geringerer Hemmung des zugeordneten Outputneurons auf ihr Eigensignal. Dadurch reagiert das Outputneuron deutlich stärker auf dieses Signal. Es meldet die Erkennung an den Cortex und erregt gleichzeitig ein angeschlossenes hemmendes Outputneuron, welches im Nucleus olivaris das Kletterfasersignal unterbricht. Somit fehlt allen übrigen Purkinjezellen, die an der gleichen Kletterfaser angeschlossen sind, die für die LTD und die LTP unbedingt notwendige tetanische Erregung durch das Kletterfasersignal. Keine andere Purkinjegruppe an dieser Kletterfaser kann nun mit dem gleichen Komplexsignal geprägt werden. Eine Mehrfachprägung wird aktiv verhindert.
Wenn nun eine Kletterfaser während ihrer Ausbreitung im Pontocerebellum mehrere hintereinanderliegende Purkinjegruppen sowie die zwischen ihnen angeordneten Golgizellen kontaktierte, konnte so viele verschiedene Komplexsignale erlernt werden, wie es Purkinjegruppen gab.
Im Pontocerebellum können in jedem Cortexcluster genauso viele verschiedene Komplexsignale erlernt werden, wie es Purkinjegruppen gibt, die an der von diesem Cortexcluster abgeleiteten Kletterfaser angeschlossen sind.
Zur Erinnerung: Aus dem Mittelwertsignal des Cortexclusters und dem davon abgeleiteten Striosomensignal wurde das Kletterfasersignal gewonnen und durch sequentielle Verteilung auf die in einer Reihe angeordneten Purkinjegruppen verteilt.
Die sequentielle Divergenz der Kletterfasersignale mag bei verschiedenen Spezies unterschiedlich stark ausgeprägt worden sein. Je mehr verschiedene Komplexsignale ein Wirbeltier erlernen konnte, umso spezifischer und angepasster wurden seine Reaktionen auf komplexe Reize. Dies verschaffte ihm einen Vorteil gegenüber anderen Arten, die dies nicht so gut vermochten.
Eine viel stärkere Zunahme der Anzahl der Purkinjezellen erfolgte durch die Signaldivergenz im Nucleus olivaris. Ein Input wurde dort (nach einer langen evolutionären Entwicklung) auf sehr viele Outputneuronen des Nucleus olivaris aufgeteilt.
Im Verlaufe der Evolution kam es im Nucleus olivaris zu einer Signaldivergenz, bei der die Anzahl der Outputneuronen zunahm. Jedes Inputneuron übertrug seine Erregung auf viele benachbarte Outputneuronen. So wurde die Anzahl der Kletterfasern, die ein und demselben Cortexcluster zugeordnet waren, schrittweise erhöht. Parallel dazu wuchs die Anzahl der Purkinjezellen im zugehörigen Cerebellumcluster.
Jedes Mittelwertsignal des Nucleus subthalamicus, welches zuvor genau eine Purkinjezelle im zugehörigen Cerebellumcluster kontaktiert hatte, teilte sich nun durch Signaldivergenz im Nucleus olivaris in dutzende, hunderte, tausende oder zehntausende Kletterfasern auf und kontaktierte eine ebenso große Anzahl von Purkinjezellen im zugehörigen Cluster. Diese Anzahl wurde zusätzlich vergrößert durch die sequentielle Divergenz, bei der jede Kletterfaser mehrere Purkinjegruppe kontaktierte.
Wenn anstelle einer Kletterfaser nun eine ganze Serie von parallel zueinander verlaufenden Kletterfasern vorhanden war, so fand auf jeder von ihnen der gleiche Prägungsalgorithmus statt. Erlernte also die erste Purkinjegruppe der Kletterfaser mit der Nummer 1 ein Prägungssignal, so lag genau das gleiche Prägungssignal auch an der Kletterfaser mit der Nummer 100 an. Daher lernte die erste Purkinjegruppe auf dieser Kletterfaser Nr. 100 genau dieses Signal. Verallgemeinert könnte man sagen: Alle ersten Purkinjegruppen auf allen Kletterfasern zum gleichen Inputneuron im Nucleus olivaris wurden gleichartig mit dem ersten Prägungssignal geprägt. Damit lag eine Mehrfachprägung in großem Ausmaß vor. Dies wäre eine enorme Verschwendung von Ressourcen.
Es musste ein neuronaler Wettbewerb organisiert werden, den nur eine Purkinjegruppe gewann.
Die Grundlage war gelegt durch die Signaldämpfung im Nucleus olivaris. Diejenigen Outputneuronen, die sehr dicht an dem Inputneuron mit dem cortikalen Mittelwertsignal angeordnet waren, wurden stärker mit Input versorgt und hatten das stärkere Kletterfasersignal innerhalb des Cerebellumclusters. Es trat eine abstandsabhängige Dämpfung auf. Sie wurden als erste mit einem Komplexsignal geprägt. Ihre Prägung fand mit zeitlichem Vorsprung statt. Die übrigen Purkinjegruppen hatten die Prägungsschwelle noch nicht erreichen können. Aber immer noch wurden mehrere Purkinjegruppen geprägt, wenn auch die Prägungsstärke unterschiedlich war.
Wenn nun dieses Komplexsignal wieder auftrat, hemmten diese bereits geprägten Purkinjezellen im Nucleus olivaris das signalliefernde Inputneuron. Dadurch wurde seine bisher tetanische Erregung gehemmt, so dass seine Erregung niederfrequent wurde.
Nun trat bei denjenigen Purkinjezellen, die mit dem ersten Signal vorgeprägt, jedoch noch nicht endgeprägt waren, ein Prozess der Entprägung statt. Dadurch wurde das teilweise erlernte Signal wieder vergessen.
Eine niederfrequente Kletterfasererregung macht die durch LTP und LTD verursachte Veränderung der synaptischen Kopplungsstärke rückgängig. Dafür ist eine niederfrequente Kletterfasererregung (z. B. von 5 Hz) für die Dauer von etwa 5 Minuten nötig. Dies zeigten Untersuchungen renommierter Neurologen.
So konnte eine Purkinjegruppe im Cluster, die zeitlich als erstes die Prägungsschwelle erreichte, bei den übrigen die durch LTP und LTD verursachten Veränderungen rückgängig machen, so dass sie wieder ungeprägte (freie) Purkinjegruppen wurden. Diese konnten nun das zweite, das dritte und jedes folgende neue Signal erlernen, indem dieser Prägungsprozess einfach für die neuen Signale wiederholt wurde.
Das niederfrequente Kletterfasersignal übernahm die Rolle eines Löschsignals. Damit wurde eine neuronale Konkurrenz zwischen der endgeprägten Purkinjegruppe und den nur teilgeprägten Purkinjegruppen realisiert, die den belegten Speicherplatz bei den teilgeprägten Purkinjegruppen wieder freigab. So konnte das Cerebellum seine Ressourcen schonend effektiv nutzen.
Der Output des Nucleus dentatus - also des Pontocerebellums - zog kopfwärts in den Cortex in ein Assoziationsgebiet und begründete dort ein sekundäres Cortexgebiet der Komplexsignale erster Stufe. Auch dieses Cortexgebiet besaß Mittelwertneuronen, die eine Clusterbildung ermöglichten. Die cortikale Projektion über die Brückenkerne und die Olive begründete im Cerebellum neue Cortexcluster zweiter Stufe, die nun Komplexsignale als Input erhielten und neue, höhere Komplexsignale erlernen konnten. Diese zogen wieder zum Cortex und begründeten Assoziationsgebiete der nächsthöheren Stufe. Dieser Prozess war rekursiv.
Nun fehlte nur noch die Möglichkeit, Komplexsignale wieder in Elementarsignale zu transformieren, damit der ursprüngliche Input - der ja im Cerebellum -verschwand- - wiederhergestellt werden konnte, da er für die bisher zugewiesenen Aufgabengebiete benötigt wurde. Dazu gab es mehrere Möglichkeiten.
Einerseits zog das Axon des Nucleus dentatus zu einem Cortexneuron der Klasse 4 und wurde dort an ein Cortexneuron der Klasse 3 übergeben. Dessen Axon zog zum motorischen Cortex und kontaktierte dort alle diejenigen Cortexneuronen, deren Elementarsignale dem Komplexsignal zugeordnet waren. So divergierte das Komplexsignal wieder auf seine Elementarsignale. Ob diese Möglichkeit tatsächlich genutzt wird, ist zurzeit noch ungewiss. Die Wahrscheinlichkeit, dass diese Divergenzvariante tatsächlich verwendet wird, ist eher unwahrscheinlich, denn auf der motorischen Seite war die Signalkonvergenz die vorherrschende Variante. Man denke nur an die Betzschen Riesenpyramidenzellen.
Eine weitere Möglichkeit ergibt sich durch eine zum Pontocerebellum inverse Schaltung, die aus den Komplexsignalen wieder die Elementarsignale herstellt. Fast ein Jahrzehnt suchte ich nach dieser inversen Schaltung und fand sie im Jahr 2012 im Cerebellum, genauer im Pontocerebellum. Ich bezeichnete diese Schaltung als inverses Cerebellum. Mehrmals musste ich setdem meine bisherigen Erkenntnisse revidieren.
Viele werden meinen, ich konstruiere hier künstlich ein kompliziertes System, nur um meine Ideen von der Arbeitsweise des Gehirns zu verbreiten. Es gibt jedoch drei Tatsachen, die unter Neurologen international anerkannt sind. Diese drei Tatsachen reichen aus, um die Existenz eines inversen Cerebellums zu beweisen. Ich zähle diese drei Tatsachen einfach auf und erkläre ihre Bedeutung.
Tatsache 1:
Das Pontocerebellum projiziert in sekundäre Cortexgebiete, die ihrerseits über die Brückenkerne zurück ins Cerebellum projizieren, wo die Signale die Körnerzellen erreichen.
Tatsache 2:
Alle cortikalen Elementarsignale projizieren in den Nucleus ruber zurück. Der Nucleus ruber seinerseits projiziert (auch) über den Nucleus olivaris in die Kletterfasern des Cerebellums.
Tatsache 3:
Es gibt im Cerebellum nur ein Verschaltungsprinzip.
Wenn man anerkennt, dass die Moosfaserprojektion und die Kletterfaserprojektion ins Pontocerebellum nach genau einem Prinzip erfolgt, es also nicht zwei oder sogar noch mehr unterschiedlich arbeitende Pontocerebellum-Bereiche gibt, dann hat man bereits die Existenz des inversen Pontocerebellums akzeptiert.
Wie ist die Arbeit des inversen Pontocerebellums zu erklären?
Der sekundäre Cortex empfängt vom Pontocerebellum die Komplexsignale und sendet sie über die Moosfaserprojektion in das inverse Cerebellum. Diese Komplexsignale sind im inversen Cerebellum der Körnerzellinput.
Der primäre Cortex sendet seine Elementarsignale zum Nucleus ruber. Dieser sendet sie zur Olive. Von der Olive gelangen diese Elementarsignale als Kletterfasersignale ins Pontocerebellum in die Abschnitte, in denen die Moosfasersignale der Komplexsignale ein eigenes Cerebellumcluster bilden. Dieses Cluster ordnen wir dem inversen Pontocerebellum zu.
Damit haben die Elementarsignale und die Komplexsignale ihre Rollen vertauscht. Stellten beim (nichtinversen) Pontocerebellum die Moosfasersignale den Output der cortikalen Elementarsignale dar, so sind im inversen Pontocerebellum den cortikalen Elementarsignale die Kletterfasern zugeordnet.
Immer, wenn ein Ensemble von Elementarsignalen aktiv ist, die im Pontocerebellum bereits einem Komplexsignal zugeordnet wurden, sind im inversen Pontocerebellum so viele Kletterfasern aktiv, wie es zugehörige Elementarsignale gibt. Denn jedes Elementarsignal, welches zum Komplexsignal gehört, ist ja aktiv und projiziert über den Nucleus ruber und die Olive ins inverse Cerebellum. Die Aktivität der Kletterfasern bewirkt über kurz oder lang durch LTP und LTD eine Prägung, bei der das aktive Komplexsignal genau diejenigen Purkinjezellen erregt, die die Elementarsignale dieses Komplexsignals repräsentieren.
Sobald danach das Komplexsignal aktiviert wird, werden im inversen Cerebellum automatisch diejenigen Purkinjezellen aktiviert, die dessen Elementarsignale repräsentieren. Ihr Output erreicht den Thalamus und dockt genau an denjenigen Neuronen an, die genau diesem Komplexsignal zugeordnet sind. Der Thalamus projiziert diese Signale wieder zum Cortex.
Ein einmal erkanntes Komplexsignal aktiviert auf diesem Wege alle ihm zugeordneten Elementarsignale im Thalamus und im Cortex. Diese aktivieren im Pontocerebellum das zugeordnete Komplexneuron, also die Purkinjezelle dieses Komplexsignals. Deren Output aktiviert im inversen Cerebellum wiederum alle zugeordneten Elementarsignale. So ergibt sich eine Signalrotation, die dieses Signal im Bewusstsein hält.
Natürlich hält diese Signalrotation nicht ewig an. Wie im limbischen System wird sie schwächer, da die Feuerrate mit der Zeit abnimmt. Und sie wird letztlich durch die laterale Hemmung unterbrochen. Sobald andere, hinreichend starke Signale den Thalamus in dem Gebiet erreichen, in dem gerade die Signalrotation stattfindet, werden diese ihrerseits hemmende Interneuronen aktivieren, die die Signaloszillation schwächen oder gar unterbrechen.
Bei der Signaloszillation zwischen Cortex, Cerebellum, inversem Cerebellum und dem Cortex kommt es zum ständigen Wechsel der Signalform. Wir unterscheiden zwei Signalformen: die Elementarform und die Komplexform.
Bei der Elementarform sind die Signalneuronen der Klasse 3 bzw. 5 des Cortex aktiv und projizieren ins Pontocerebellum. Das zugeordnete Komplexneuron - eine Purkinjezelle bzw. die Purkinjezellen einer Purkinjegruppe - erkennen dieses Komplexsignal und antworten mit einem Output. Dieser Output liegt in der Komplexform vor. Er erreicht das sekundäre Cortexgebiet und projiziert von dort in die Moosfasern des inversen Cerebellums, während die Elementarsignale ins Kletterfasersystem dieses Gebietes projizieren. Die Moosfasern erregen diese Purkinjezellen, deren Output wieder genau die Elementarsignale sind. Damit ist der Wechsel von der Komplexform in die Elementarform vollzogen und die Oszillation beginnt von vorn.
Diese Signaloszillation wird real im menschlichen Gehirn beobachtet.
Welchen Nutzen bringt sie für den Menschen?
Ein Nutzen besteht in der größeren Lebensdauer eines Komplexsignals. Es wird in den Rotationsschleifen sozusagen zwischengespeichert. So mancher Gedanke lässt sich einfach nicht verdrängen.
Ein viel größerer Nutzen liegt darin, dass nun im Gehirn eine automatische Vervollständigung von unvollständigen Signalen möglich wird.
Wir alle kennen Kreuzworträtsel. Es gilt, Buchstaben sind zu finden, zu denen es gewisse Hinweise gibt. Mit der Zeit füllen immer mehr Buchstaben die Zeilen und Spalten des Rätsels, dennoch bleiben vor allem anfangs noch viele Buchstabenlücken. Der Mensch ist in der Lage, zu lückenhaft vorliegenden Wörtern die fehlenden Buchstaben zu erkennen.
Wie funktioniert das?
Wenn ein komplettes Wort im Pontocerebellum in einer Purkinjegruppe abgespeichert ist, während die einzelnen Buchstaben - jeder für sich - auf einer Moosfaser die Purkinjezellen eines zugeordneten -Spracherkennungsclusters- im Pontocerebellum erregen, so wird beim Wort -Fahrrad-, welches unvollständig in Form von -F hr ad- vorliegt, jede Purkinjezelle aktiviert, die die Moosfasern der Teilbuchstaben f,h,r,a und d anzapft und von ihnen aktiviert wird. Die dem Wort Fahrrad zugeordnete Purkinjezelle wird das Wort erkennen. Ihr Output ist jedoch schwächer, als wenn das komplette Wort vorliegen würde. Jeder fehlende Buchstabe vermindert den Output des zuständigen Dentatusneurons.
Gibt es viele Wörter mit den Teilbuchstaben f,h,r, a und d, so gibt es mehrere Lösungen. Dann sind gleichzeitig mehrere Purkinjezellen aktiv und melden mehrere Lösungen an den Cortex. Je nach Übereinstimmung mit unterschiedlicher Stärke. In unserem Fall gibt es jedoch kaum andere Wörter als das Wort Fahrrad. Die Lösung ist eindeutig.
Das Cerebellumsystem kann also unvollständige Signale vervollständigen.
Wir erkennen ein Pferd, auch wenn wir es nicht komplett sehen. Die Frage -Wer frisst Hafer- reicht aus, um das Pferd zu nennen. Aber ebenso andere Tiere, die Hafer fressen. Und ganz abstrakt könnte man sogar vermuten, ein Mähdrescher fresse Hafer, weil dieser in ihm verschwindet. Eine Teilinformation genügt oft, um die fehlende Information zu erkennen.
Fasst man unvollständige Signale als Fragen auf, so liefert das Cerebellumsystem die Antwort. Aus dem Unterbewusstsein entspringt die Antwort: -Ein Pferd frisst Hafer.- Und nun hat dieses Unterbewusstsein einen Namen und eine materielle Repräsentation im Gehirn. Es ist das Pontocerebellum, welches die Antwort liefert. Genauer das inverse Cerebellum, welches die Signalvervollständigung bewirkt.
Wenn man beachtet, dass die sekundären Cortexgebiete, die vom Cerebellum die Komplexsignale empfangen, auch nur gewöhnliche Cortexgebiete sind, kann man den weiteren Werdegang vorhersehen. Die Komplexsignale aktivieren Cortexneuronen, die ihrerseits wieder als Elementarsignale aufgefasst werden können. Denn ein Elementarsignal liegt genau dann vor, wenn es durch die Aktivität eines einzigen Cortexneurons repräsentiert wird. So sind die Komplexsignale erster Stufe praktisch Elementarsignale zweiter Stufe. Sie projizieren wieder über Brückenkerne und die Moosfasern in die Körnerzellen eines neuen Clusters im Pontocerebellum. Dieses könnte man auch als Cluster zweiter Stufe bezeichnen.
Ebenso bilden in diesem sekundären Cortexgebiet Mittelwertneuronen der Klasse 6 die Mittelwertsignale, die ihrerseits auf Neuronen im Nucleus subthalamicus konvergieren. Über den Nucleus ruber erreichen sie die Olive und das Pontocerebellum. So erhält das Cerebellumcluster der Stufe 2 seine Kletterfaserprojektion. Der Lernprozess - die Prägung von neuen Komplexsignalen der zweiten Stufe - kann beginnen.
Dieser Prozess ist rekursiv. Komplexsignale der Stufe k werden in einem neuen Cortexgebiet der Stufe k zu Elementarsignalen der Stufe k+1. Diese bewirken im Pontocerebellum die Bildung neuer Komplexsignale der Stufe k+1. So geht es weiter, bis die Anzahl der beteiligten Körnerzellen, Golgizellen, Stern- und Korbzellen und vor allem der Purkinjezellen aufgebraucht ist.
Und in jeder dieser Stufen entsteht im Pontocerebellum ein zugeordnetes inverses Cerebellum, welches in der Lage ist, die neuen Komplexsignale aus der Komplexform wieder in die Elementarform zu transformieren. In jeder Ebene findet eine Signalrotation statt, bei der die Signalform ständig oszilliert zwischen der Komplexform und der Elementarform des jeweiligen Signals.
Hier verselbständigt sich dieser Prozess.
Auf den unteren Stufen - beispielsweise im primären Cortex - treffen die Elementarsignale des inversen Cerebellums im Thalamus ein und werden meist von den aktuell eintreffenden Signalen durch Hemmung unterdrückt. Dies liegt auch an der engen Nachbarschaft dieser Signale.
Bei Komplexsignalen der höheren Stufen ist dies nicht mehr der Fall. Da bei ihnen deutlich mehr Elementarneuronen beteiligt sind, die demzufolge auch auf eine größere Fläche im Thalamus verteilt sind, besitzen sie untereinander größere Abstände. Dadurch wird die Nachbarhemmung geringer. Sind sogar verschiedene Modalitäten im Komplexsignal miteinander gekoppelt, ist eine laterale Hemmung unmöglich. Ab einer gewissen Stufe ist der Abstand der Elementarneuronen so groß, dass faktisch keine laterale Nachbarhemmung auftritt. Dann wird die Signalrotation nicht mehr unterbrochen. Praktisch findet die nötige Unterbrechung der Signalrotation dann über die Hemmung des Thalamus statt, die vom circadianen Rhythmus des Nucleus suprachiasmaticus gesteuert wird und während des Schlafes stattfindet.
Diese rotierenden Komplexsignale sind uns ständig bewusst. Und sie liegen gleichzeitig in der Elementarform vor, so dass wir instinktiv wissen, wie das Pferd aussieht, welches den Hafer frisst. Und wir wissen, wie es riecht, wie es läuft, galoppiert, wie es wiehert. Nicht nur der abstrakte Begriff -Pferd- rotiert in unserem Kopf, sondern auch - auf tieferen Stufen - sämtliche uns bekannten Eigenschaften. Dies ist offenbar ein Merkmal unseres Bewusstseins.
Hier zeigt sich die große Schwäche des Human Brain Projekts. Menschen, die noch nie ein Pferd gesehen haben, auch nie von einem Pferd hörten, werden die synaptischen Verbindungen des Komplexsignals -Pferd- nicht im Pontocerebellum und im inversen Pontocerebellum, im Thalamus und Cortex besitzen. Genau diese synaptischen Verbindungen haben jedoch diejenigen, die Pferde kennen.
Doch während der eine meint, ein Pferd pflüge den Acker des Bauern, meint ein anderer, Pferde seien Säugetiere, ein Dritter denkt beim Pferd nur ans Reiten. So sind die synaptischen Verbindungen dieser Personen zum Thema Pferd völlig verschieden. Speziell die Assoziationsgebiete der Gehirne verschiedener Menschen unterschieden sich in der konkreten Art der synaptischen Verknüpfungen. So stark, wie sich ihr Kulturkreis, ihr Bildungsstand, ihre Interessen, ihre Muttersprachen und die von ihnen in Komplexsignale umgesetzten Erkenntnisse voneinander unterscheiden, so stark unterschieden sich die synaptischen Verbindungen in den höheren Assoziationsgebieten des Cortex, in den Clustern des Pontocerebellums, in der Matrix der Basalganglien, in den Rotationsschleifen des limbischen Systems und in vielen anderen Substrukturen, die es noch zu erkennen gilt.
Lediglich die primären Cortexgebiete, die etwa visuelle Divergenzmodule zum Farbsehen, Richtungssehen usw. enthalten, sind gleichartig aufgebaut. Doch auch hier gibt es Unterschiede in der Schichtdicke, der Anzahl der Inputneuronen und Outputneuronen sowie im konkreten Aufbau der inversen Module. Daher ist der virtuelle Nachbau eines Gehirns mit all seinen Synapsen immer nur der Nachbau eines speziellen Menschengehirns, welches personengebunden ist. Ob der riesige Arbeitsaufwand lohnt und auf andere Gehirne übertragbar ist, sei kritisch hinterfragt.
Viel erfolgversprechender ist die hier beschriebene Herangehensweise. Sie kann erklären, wie Gedanken, Gefühle, Vorstellungen und Bewusstsein im Gehirn entstehen kann.
Bereits in meiner ersten Monografie formulierte ich diese Erkenntnis.
Theorem über die Entstehung von Bewusstsein im Gehirn
Mit der wachsenden Anzahl der Signale im rekursiv aufgebauten System aus den Cerebellumclustern des direkten und inversen Pontocerebellums nimmt die Hemmungsstärke in der rezeptiven Nachbarhemmung des Thalamus mit zunehmender Stufenzahl der Rekursionen abstandsbedingt ab.
Dadurch kann sich der Output der inversen Systeme verselbständigen und in Überlagerung und Rückkopplung mit dem aktuellen sowie mit dem früheren, zwischengespeicherten Input ein inneres, multimodales und zeitveränderliches Abbild der Welt erschaffen, welches als Bewusstsein bezeichnet werden kann.
Hierbei treten im Gehirn drei Signalstufen auf, die drei Subsysteme bilden. Jedes Subsystem besteht aus einem aufsteigend projizierenden Modul und einem absteigend projizierenden Modul, welches invers zum aufsteigenden arbeitet. Auf der untersten Stufe liegen die Signale in der analogen Form vor. Jedem analogen Signal ist, wenn es aktiv ist, eine Feuerrate zugeordnet. Diese Stufe bezeichnen wir hier als primäres Subsystem.
Im sekundären System liegen die Signale in extremwertcodierter Form vor. Eine Neuronenpopulation aktiver Neuronen ist gekennzeichnet durch ein lokales Erregungsmaximum, welches die Parameter verschlüsselt.
Im dritten, tertiären System liegen alle Signale in der Komplexform vor. Ausführlicher wird dies im letzten Kapitel dargelegt.
Für Anhänger der neuronalen Netze sei hier kurz angemerkt, dass ein Cluster im Pontocerebellum vereinfacht eine Assoziativmatrix ist. Die waagerechten Zeilen entsprechen den Moosfasern, die senkrechten Zeilen den Purkinjezellen. Die Kreuzungspunkte stellen die Synapsen dar. Der Output des Clusters speist eine zweite Assoziativmatrix, bei der jedoch Input und Output vertauscht sind: Die Outputsignale der ersten Assoziativmatrix speisen die waagerecht verlaufenden Zeilen der zweiten Assoziativmatrix, während die senkrechten Spalten der zweiten Assoziativmatrix ihren Input von den waagerecht verlaufenden Zeilen beziehen.
Dies stellt eine Rückkopplung dar, bei der jedes in der ersten Matrix erkannte Signal als Input für die zweite Matrix verwendet wird und durch sie wieder in seine ursprünglichen Elementarsignale zerlegt wird. So findet ebenfalls eine Signalrotation statt.
In diesem vereinfachten Modell fehlt nur noch die Einbindung der Kletterfasersignale derart, dass der in neuronalen Netzen übliche und langwierige Prozess der -Prägung- durch mittelwertgesteuerte Schreibimpulse beschleunigt wird. Eine Zielfunktion zur Steuerung des Prägungsprozesses ist dann völlig überflüssig.
8.1 Die
Speicherpyramide des Cerebellums
Bisher hatten wir die Speicherung von Signalen am
Beispiel einer cortikalen Clustergruppe betrachtet (Abbildung 40). Nun
besteht der Cortex aus unzählig vielen derartigen Clustergruppen.
Wir unterteilen also den gesamten primären Cortex
in Cluster. Das Hauptmerkmal eines Clusters besteht in einem gemeinsamen
Mittelwertneuron der Klasse 6, zu jedem gibt es ein derartiges Cluster. Wir
denken uns (vereinfacht) jedes Cluster als kleines Quadrat mit vielen
tausend Outputneuronen (Elementarneuronen).
Nun bilden wir so viele Clustergruppen wie
möglich in dieser cortikalen Fläche. Jede Clustergruppe besitzt ein
Mittelcluster und 8 Randcluster. Ihr ist im Cerebellum genau ein
Cerebellumcluster zugeordnet. Dieses Cerebellumcluster erlernt im Verlauf
des Lebens des Lebewesens unzählige Komplexsignale, deren Elementarsignale
genau aus der zugeordneten cortikalen Clustergruppe kommen.
Jedes der vielen Cerebellumcluster projiziert für
jedes erlernte Komplexsignale in ein neues, sekundäres Cortexgebiet. somit
entspricht jedem primären Cortexcluster genau ein sekundäres Cortexcluster.
Die sekundären Cortexcluster sind Gebiete, in denen jedem Elementarneuron
genau ein Komplexsignal des primären Cortex entspricht. Der Gesamtheit der
sekundären Cortexcluser entpricht eine (meist) zusammenhängende Cortexfläche
im sekundären Cortex.
Nun beginnt der Lernprozess erneut von vorn. Auch
der sekundäre Cortex besitzt Mittelwertneuronen und Elementarneuronen. Die
Mittelwertneuronen projizieren über den Nucleus ruber und den Nucleus
olivaris in die Kletterfasern eines neuen Cerebellumgebietes im
Pontocerebellum. Dorthin projizieren auch die Elementarneuronen der
sekundären Cortexrinde. So kann dieses neue Cerebellumgebiet wieder neue
Complexsignale erlernen. Diese bestehen dieses mal jedoch aus der
Kombination von Komplexsignalen, denn sie entstammen ja bereits dem
Pontocerebellum.
So erlernt das Pontocerebellum Kompexsignale
zweiter Stufe.
Da diese wieder über eine cortikale Projektion im
Cortex, einem tertiaren Cortex, also dem Cortex dritter Stufe landen, können
sie unter Zuhilfenahme der Cerebellumprojektion über die Kletter- und
Parallelfasern neue Komplexsignale dritter Stufe erlernen.
Das ganze kann sich so oft wiederholen, bis es im
Cerebellum keine freien Gebiete mehr gibt, die neue Signale erlernen können.
Dies ist durch die Obergrenze der möglichen Purkinjezellen begrenzt.
Irgendwann könnte (oder wird tatsächlich) die Zahl der verfügbaren
Purkinjezellen ausgeschöpft sein.
Doch dieses System, das man sich durchaus als
Speicherpyramide des Cerebellums vorstellen kann, ist der Grund dafür, dass
mit zunehmender Komplexität der Signalauswertung immer komplexere
Komplexsignale erlernt und miteinander kombiniert werden können.
Erwähnt werden sollte an dieser Stelle auch, dass
alle Cortexgebiete, also die primären, sekunderen, tertiären uns solche von
noch höherer Stufe ebenfalls ins Basalgangliensystem und in das limbische
System projizieren, so dass nicht nur Komplexsignale der aktuellen
Gegenwart, sondern auch die der unmittelbaren Vergangenheit (Basalganglien)
und der länger zurückliegender Vergangenheit (limbisches System) in die
Bildung neuer Komplexsignale einbezogen werden können.
Die neuronale Verknüpfungsstruktur der höheren
Cortex- und Cerebellumgebiete ist also davon abhängig, wann und in welcher
Reihenfolge welche Komplexsignale (z. B. Sprache) erlernt werden. Daher ist
die Verknüpfungsstruktur bei allen Menschen verschieden voneinander. Und wer
diese neuronale Verknüpfungsstruktur bis ins letzte Detail bei einem
bestimmten Gehirn aufdeckt, kann nicht behaupten, die Verknüpfungsstruktur
der Gehirne aller Menschen entschlüsselt zu haben.
Lediglich das Verknüpfungsprinzip, welches auf
der Clusterbildung von Cortex und Cerebellum beruht, ist bei den
Wirbeltieren und den Menschen einheitlich.
Offenbar hat der Mensch bezüglich der
Speicherpyramide des Cerebellums die höhste Stufenanzahl erreicht. Nicht zu
vergessen ist jedoch der Einspareffekt an Neuronen durch die vom Menschen
entwickelten Komprimierungsfähigkeiten. Sprache – und auch begriffliches,
teils symbolisches Denken – ermöglichen eine extrem starke
Datenkomprimierung. Man vergegenwärtige sich, dass ein Film von etlichen
Megabyte Signalinhalt beispielsweise durch ein Drehbuch mit wenigen hundert
Seiten oder durch eine Kurzbeschreibung in einer TV-Programmzeitschrift
relativ gut wiedergegeben werden kann. Ohne Datenkomprimierung wäre der
Mensch geistig nicht sonderlich von seinen Vorfahren zu unterscheiden.
Inwieweit sich die Gehirnstruktur des Menschen
von anderen Wirbeltieren unterscheidet, ist noch Gegenstand der Forschung.
Jedoch ist davon auszugehen, dass die meisten Gehirnstrukturen in allen
Wirbeltieren hervorgebracht wurden und ihre Leistungsfähigkeit im Verlauf
der Evolution verbessert wurde. Der Mensch hatte als Spezies einfach mehr
Glück.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan