Gehirntheorie des Menschen
ISBN 978-3-00-068559-0
Monografie von Dr. rer. nat. Andreas Heinrich Malczan
9. Das inverse Pontocerebellum
Bereits im Jahr 2012 entwickelte ich das Konzept des inversen Cerebellums. Ein Grund dafür war das von mir entdeckte Prinzip, dass jegliche neue Signalform, die durch irgendein Modul hervorgebracht wird, immer in die ursprüngliche Signalform zurücktransformiert werden muss, damit sie ihre bisherigen Aufgaben im System wahrnehmen kann.
Wenn nun das Pontocerebellum die Elementarsignale in Komplexsignale überführt, so muss ein weiteres Modul diese Komplexsignale wieder in die ursprünglichen Elementarsignale zurücktransformieren. Sonst würden diese im Pontocerebellum einfach für immer verloren gehen, anstatt ihre bisherigen Wirkungen zu verursachen.
Da das Komplexsignal zu einer aktiven Neuronenpopulation von Elementarsignalen vom Pontocerebellum gebildet wird, erhielt die theoretisch notwendige inverse Struktur zunächst den Namen „inverses Cerebellum“.
Die Funktionsweise war prinzipiell klar. Das Pontocerebellum bildete aus Elementarsignalen die neue Klasse der Komplexsignale. Hierzu benötigte es die Mittelwert-Kletterfasersignale für die Prägung der Purkinjezellen mittels LTD und LTP. Die Elementarsignale waren der Moosfaserinput. Der Output des Pontocerebellums waren die Komplexsignale.
Diese mussten im inversen Pontocerebellum wieder in Elementarsignale transformiert werden.
Die ursprünglichen Elementarsignale zogen vom (primären) Cortex (unter anderem) zum Nucleus ruber. So erreichten sie (auch) den Seitenwechselkern dieser Etage, den Nucleus olivaris. Von dort zogen sie als Kletterfasern zum Pontocerebellum.
Nun ist für den Prägungsprozess ein getaktetes Kletterfasersignal erforderlich, es muss also durch kurze Pausen unterbrochen werden. In diesen Pausen erkennen Purkinjegruppen ihr Eigensignal und hemmen die Kletterfasersignale der Olive, damit keine Mehrfachprägung auftritt.
Die kurzen Pausen bewerkstelligte das Basalgangliensystem. Es empfing ebenfalls die gleichen Elementarsignale, bewirkte eine kurze Zeitverzögerung und projizierte hemmend in den (ventralen) Thalamus, wo sie die ursprünglichen Elementarsignale unterbrachen. Da dadurch auch der Signalfluss zum Cortex und somit auch zu den Basalganglien unterbrochen wurde, fiel mit einer gewissen Zeitverzögerung die Hemmung der Elementarsignale weg. Dies alles galt jedoch nur für stationäre Signale, also solche, die eine hinreichend lange Signaldauer besaßen. Für kurzlebige Signale bewirkten die Basalganglien die Bewegungserkennung, indem das zeitverzögerte Echo hemmend wirkte, so dass von unveränderten Signalen kein Restsignal übrig blieb. So wurden Bewegungen und Signalveränderungen erkennbar.
Bei stationären (unveränderlichen Signalen mit einer längeren Signaldauer) war es anders. Hier wurde den Elementarsignalen die kurzen Pausen aufgezwungen, die für die Prägung im Pontocerebellum unbedingt nötig waren.
Jedes Elementarsignal aktivierte so nicht nur die Moosfasern im Pontocerebellum, sondern auch (mindestens) eine Kletterfaser im inversen Pontocerebellum. Wir erinnern uns daran, dass (nach dieser Theorie) jedes cortikale Elementarsignal bereits in der Frühzeit über den Nucleus ruber und den Nucleus olivaris zur Gegenseite zog und dort als Kletterfaser das Cerebellum erreichte. Nun brauchte es nur noch weiterzuziehen und im Mittelwertgebiet, dem Pontocerebellum, eigene Purkinjezellen zu generieren.
Durch Prägung erlernten diese Purkinjezellen nun alle Komplexsignale, die während der Aktivität dieses Elementarsignals aktiv waren. Durch eine Signaldivergenz der Kletterfasern (im Nucleus olivaris) konnten auch Purkinjezellen in unterschiedlichen Segmenten des inversen Pontocerebellums Komplexsignale in ihre Prägung mit einbeziehen, die genau dieses Elementarsignal in ihrem Signalspektrum aufwiesen. Denn viele Elementarsignale kamen in verschiedenen Komplexsignalen als Signalbestandteil vor.
Immer, wenn ein Komplexsignal aktiv war, zu dessen Prägungsmenge ein bestimmtes Elementarsignal gehörte, wurde genau die Purkinjezelle im inversen Cerebellum aktiviert, die diesem Elementarsignal zugeordnet war.
So überführte das inverse Pontocerebellum alle Komplexsignale wieder in die zugeordneten Elementarsignale, wobei hierfür die Prägung, also ein Lernprozess nötig war. Erst nach dessen Abschluss konnte ein Komplexsignal wieder in die Elementarsignale überführt werden.
Der Output des inversen Pontocerebellums nahm den Weg zum Thalamus und kontaktierte dort genau diejenigen Neuronen, die den Elementarsignalen zugeordnet waren.
Führte nun die Aktivität einer bestimmten Neuronenpopulation im Thalamus zur Erregung der zugehörigen Elementarneuronen im Cortex, so erreichten diese Signale über die Brückenkerne und die Moosfasern die zugeordnete Purkinjezelle. Diese erkannte dieses Signal als ihr Eigensignal und aktivierte im sekundären Cortex ein Komplexneuron.
Dieses Komplexneuron aktiviert im inversen Pontocerebellum eine zugeordnete Population von Purkinjegruppen, von denen jede einem der aktiven Elementarsignale zugeordnet ist. Der Output des inversen Pontocerebellums aktiviert nun im Thalamus genau die Neuronenpopulation, die diesem Komplexsignal zugeordnet ist.
So entsteht eine Rückkopplung, die das aktive Komplexsignal und die ihm zugeordneten Elementarsignale stetig neu aktiviert. Das Signal oszilliert mit einem ständigen Wechsel seiner Signalform von der Elementarform zur Komplexform und wieder zurück zur Elementarform.
Diese Signaloszillation wurde im menschlichen Gehirn real nachgewiesen (z. B. von Wolf Singer).
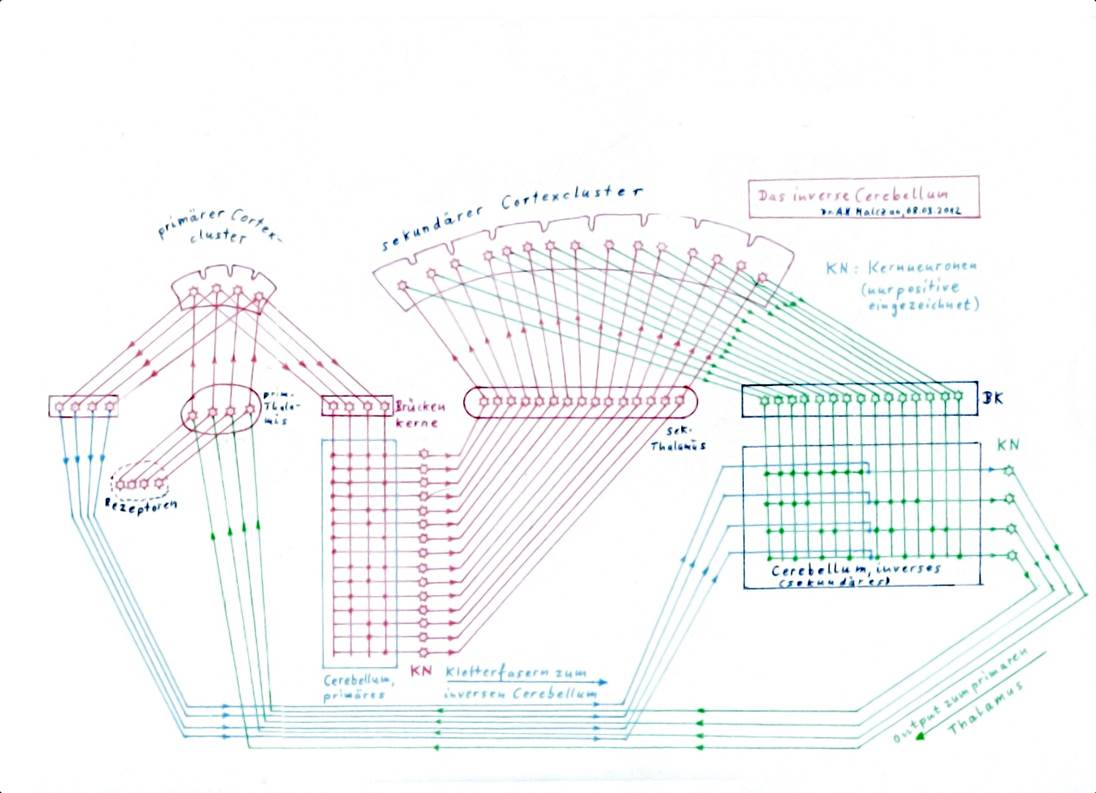
Abbildung 48: Cerebellum und inverses Cerebellum - Grundschaltung
In der obigen Abbildung ist der Weg der cortikalen Elementarsignale zum inversen Cerebellum stark vereinfacht. In Wirklichkeit besteht er aus mehreren Etappen und zwei parallelen Wegen. Der Cortex projiziert über den Nucleus ruber zum Nucleus olivaris, der seinerseits die Kletterfasern zum inversen Cerebellum sendet (oben grün dargestellt). Andererseits projiziert der Cortex in die Substantia nigra, die zurückprojiziert zum Striatum, welches wiederum hemmend in den Thalamus projiziert und dort bei stationären Signalen eine kurzzeitige Signalunterbrechung bewirkt. Diese Signalunterbrechung wird über den Cortex an die Kletterfaserprojektion zum inversen Cerebellum weitergegeben. So erhalten die Kletterfasersignale auch im inversen Cerebellum ihre typische Form, eine Schwingung, die von kurzen Pausen unterbrochen wird. Sie ist für die Prägung von Purkinjezellen bestens geeignet.
Welchen Sinn sollte es haben, wenn ein Komplexsignal alle seine Elementarsignal-Komponenten aktiviert? Schließlich sind die ihm zugeordneten Elementarsignale ja sowieso aktiv, wenn das Komplexsignal aktiv ist.
Der Sinn ergibt sich dadurch, dass im Pontocerebellum auch unvollständige Signale erkannt werden. Gehören zu einem bestimmten Komplexsignal beispielsweise 200 Elementarsignale, so liefert das zugehörige Dentatusneuron auch dann ein Outputsignal, wenn z. B. nur 150, 120 oder gar nur 100 von diesen 200 Elementarsignalen aktiv sind.
Das Dentatusneuron meldet dann dem sekundären Cortex die Erkennung dieses Komplexsignals, obwohl ja nur knapp die Hälfte der nötigen Elementarsignale aktiv war. Wenn nun das inverse Pontocerebellum dieses Komplexsignal als Input erhält und daraus alle zugeordneten Elementarsignale komplett aktiviert, so sind neben den 150, 120 oder gar nur 100 bereits aktiven Elementarsignalen plötzlich alle 200 Elementarsignale aktiviert. Dadurch sind nun plötzlich 50, 80 oder gar 100 Elementarsignale zusätzlich aktiv, die zuvor völlig inaktiv waren.
So ergänzt das inverse Cerebellum unvollständige Signale auf der Ebene der Elementarsignale. Diese Signalergänzung stellt den vom Gehirn selbsterschaffenen Input dar, den wir – auf höheren Stufen – durchaus als Gedanken, Eingebungen, Ideen usw. betrachten können. Sie entsteigen dem Unterbewusstsein, dessen materieller Sitz im Cerebellum verortet werden muss, und erreichen das Bewusstsein im Cortex.
Wird die Frage „Wer frisst Hafer“ gestellt, so ergänzt das inverse Pontocerebellum die Antwort, weil das Komplexsignal „Pferde fressen Hafer“ aktiviert wird, auch wenn es nur unvollständig ist. Es wird aktiviert, weil es die Teilsignale „Hafer“ und „fressen“ besitzt. Lediglich das Teilsignal „Hafer“ fehlt. Dennoch reicht es für die Aktivierung dieses Komplexsignals. Das Komplexsignal „Pferde fressen Hafer“ seinerseits aktiviert im inversen Pontocerebellum die Signalbestandteile (Elementarsignale) „Pferde“, „Hafer“ sowie „fressen“. Da die Signale „Hafer“ und „fressen“ bereits Bestandteile der Frage waren, sind sie aktiv und unterdrücken den Output des inversen Pontocerebellums. So bleibt als Antwort nur das Signal “Pferde“ übrig. Damit ist die Antwort gefunden: „Wer frisst Hafer? Pferde!“
Ohne inverses Pontocerebellum blieben die Komplexsignale des Pontocerebellums in diesem stecken, es gäbe keinen Algorithmus, der selbsterschaffene Signale produziert. Insofern verdanken wir unser Denken vorwiegend – jedoch nicht nur – dem Zusammenspiel von Pontocerebellum und inversem Pontocerebellum.
Möglich wird dies alles durch die Rückkopplung zwischen dem Pontocerebellum und dem inversen Pontocerebellum. Und da man beide durchaus als (modifizierte) neuronale Netze interpretieren kann, werden sich die Anhänger der rückgekoppelten neuronalen Netze bestätigt fühlen in ihrer Annahme, dass nur eine sinnvolle Rückkopplung künstliche Intelligenz hervorbringen kann, die mehr kann als nur Mustererkennung.
Bereits in meiner ersten Monografie formulierte ich diese Erkenntnis:
Theorem über die Entstehung von Bewusstsein im Gehirn
Mit der wachsenden Anzahl der Signale im rekursiv aufgebauten System aus den Cerebellumclustern des direkten und des inversen Pontocerebellums nimmt die Hemmungsstärke in der rezeptiven Nachbarhemmung des Thalamus mit zunehmender Stufenzahl der Rekursionen abstandsbedingt ab.
Dadurch kann sich der Output der inversen Systeme verselbständigen und in Überlagerung und Rückkopplung mit dem aktuellen sowie mit dem früheren, zwischengespeicherten Input ein inneres, multimodales und zeitveränderliches Abbild der Welt erschaffen, welches als Bewusstsein bezeichnet werden kann.
Der Unterschied zwischen klassischen neuronalen Netzen und dem Cerebellum ist die Zweiteilung des Inputs: Die Elementarsignale kommen über das Moosfasersystem, sie sind der eigentliche Input des Pontocerebellums. Die Mittelwertsignale für das Pontocerebellum bzw. die Elementarsignale für das inversen Pontocerebellum sind die Schreibsignale, also die Schreibbefehle, die zum Abspeichern der aktuellen Signalkombination der Datenleitungen in den Purkinjezellen führen. Diese Schreibbefehle gibt es in klassischen neuronalen Netzen nicht, obwohl Google inzwischen die Prägung der künstlichen Neuronen im neuronalen Netz auch mit bestimmten Algorithmen beschleunigt. Und ein niederfrequentes Schreibsignal, das dann als Löschsignal wirkt, existiert in den klassischen neuronalen Netzen nicht.
Eine Zielfunktion und einen Abgleich mit ihr gibt es im Cerebellum ebenfalls nicht. Dennoch ist das System selbstlernend. Wird es mit „schwachsinnigem“ Input gefüttert, so kommt auch Schwachsinn heraus. Beispiele sind die Anhänger von Verschwörungstheorien, die in eigenen Informationsblasen leben und sich selbst in ihren Ansichten bestärken.
Ein Großteil des erlernten Wissens wird auch nicht mehr durch eigene „Erfahrung“ erlernt, sondern über Unterricht weitergegeben und aus vielen Quellen erworben. Insofern wäre eine künstliche Intelligenz auch diesen Einflüssen ausgesetzt und könnte leicht missbraucht werden. Völlig unerforscht ist übrigens die Entstehung eines eigenen Willens in solchen künstlichen Systemen, die dann nicht mehr extern steuerbar wären.
Es gibt im Gehirn des Menschen außer dem Cerebellum noch andere Systemkomponenten und ihre dazu inversen Subsysteme, die zur Intelligenz beitragen, die aber in dieser Monografie nicht vorgestellt werden. Grund sind die mangelnden Reaktionen auf meine bisherigen Bemühungen, die Arbeitsweise des Gehirns auf solide wissenschaftliche Füße zu stellen. Es scheint, das große Interesse an der Arbeitsweise des menschlichen Gehirns ist offenbar häufiger nur vorgetäuscht, um großzügige Fördermittel zu erhalten und gleichzeitig die wissenschaftliche Konkurrenz kleinzuhalten. Außerdem scheint es auch Dogmen zu geben, für deren Verteidigung man auch bereits ist, die Freiheit der Forschung zu opfern. Wer gegen den Strom schwimmt, mag Recht haben, aber es wird ihm wenig nützen. Oft müssen Anhänger einer gängigen Ansicht erst aussterben, bevor man neue Gedankenwege zulässt. Eine solche Ansicht ist die, dass man die Arbeitsweise des Gehirns nicht erkennen könne. Obwohl unbewiesen, scheint man zu glauben, dass das Gehirn sich nicht selbst erkennen könne. Hierbei vergisst man, dass das sich selbst erkennenden Gehirn nicht aus sich selbst heraus zu seiner eigenen Erkennung fähig ist, sondern dass es das in Jahrtausenden erworbene Wissen von hunderttausenden Wissenschaftlern lediglich anwendet, um Zusammenhänge zu erkennen, die man bisher vernachlässigt hat. Die Beachtung der nichtlinearen Signaldämpfung von neuronalen Signalen und die daraus entstehenden Konsequenzen für die Überlagerung solcher Signale würde den Verantwortlichen des Human Brain Projekts ein Mittel in die Hand geben, das Prinzip der Extremwertcodierung im menschlichen Gehirn und im Wirbeltiergehirn zu verstehen und damit einen neuen Zugang zum Verständnis seiner Arbeitsweise eröffnen. Mögen meine Worte nicht ungehört verhallen!
Monografie von Dr. rer. nat. Andreas Heinrich Malczan