Human brain theory
ISBN 978-3-00-068559-0
Monograph of Dr. rer. nat. Andreas Heinrich Malczan
10 The module of the rotational memory in the limbic system
Already in my monograph "The Limbic System" ISBN 978-3-00-037458-6 (German) or ISBN 978-3-00-042153-2 (English) I explained the limbic system and outlined a hypothesis about its origin. The uppermost segment of the ancient rope ladder system was the olfactory. The signals from the olfactory receptors arrived at the sensory centre of the first segment. Via class 3 neurons, they travelled to the motor centre, where they activated class 5 neurons, which in turn activated the body's motor neurons. Thus olfactory signals were used to control the body to find food. But the olfactory signals evaporated quite quickly in the water - where all animals lived at that time. Then the olfactory signal was gone.It was favourable to extend the duration of the olfactory signal. Here nature found the following solution.
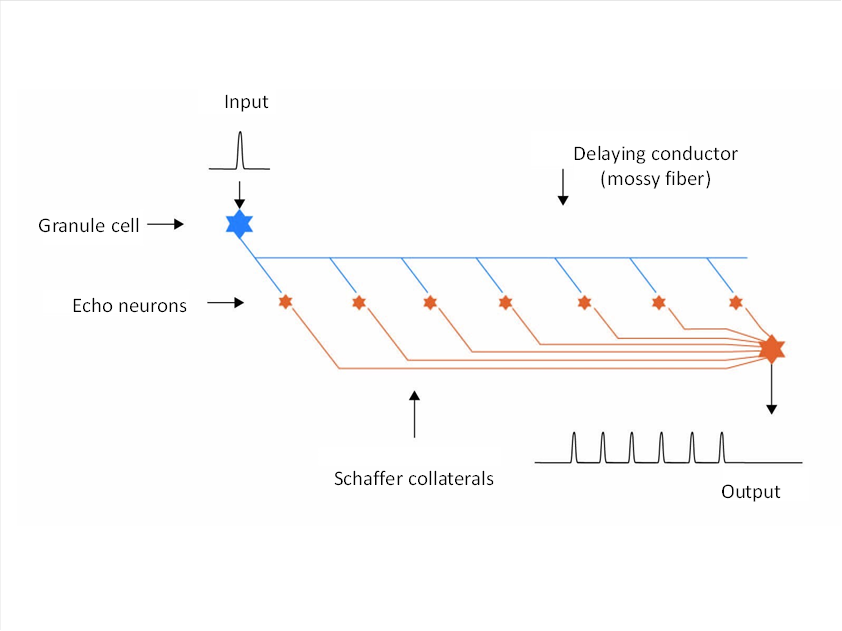
Figure 49: Basic structure of the hippocampus using the example of a moss fibre
The class 3 neurons each formed an axon, the so-called moss fibre, which formed axon collaterals at regular intervals, which in turn generated new class 3 neurons. These neurons were echoneurons that were activated by the passing action potential on the mossy fibre and thus formed an action potential themselves. Since the moss fibres had no myelin sheath, they propagated the action potentials relatively slowly. The axons of the echoneurons - the Schaffer collaterals - ended on a separate output neuron for each moss fibre. Thus, each action potential of an olfactory receptor was converted into a sequence of action potentials that now lasted much longer. The length of the moss fibres increased, as this was advantageous. This allowed the animal to react longer to olfactory stimuli, even if they only lasted a short time.
The structure that emerged was the hippocampus. Its output reached the motor nucleus, the amygdala, on the motor side. When this projected back to the hippocampus, a closed signal loop was created.
In this, an olfactory signal once generated could rotate practically indefinitely. This made a longer prey search possible. Once the prey had been eaten, an inhibitory signal from the digestive organs in the amygdala could activate inhibitory interneurons that stopped the eternal rotation of the olfactory signal.
Other substructures of the nervous system could also control the inhibitory neurons in the amygdala and stop the signal rotation. On the other hand, equally excitatory signals could trigger a new signal rotation, so that a rotation memory was now available for signals of all kinds.
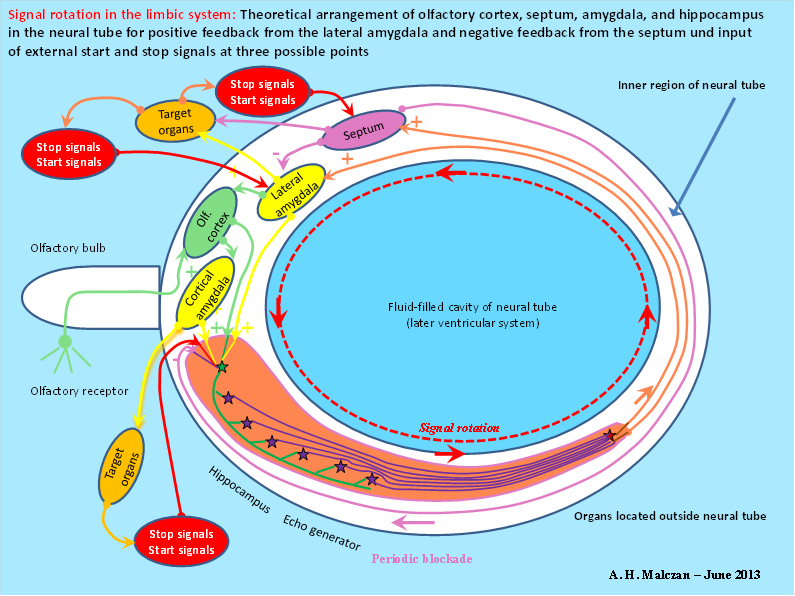
Figure 50: Principle of signal rotation in the limbic system according to A. Malczan, 2013
The septum represented the median nucleus in the first floor of the cord ladder system. Its neurons already used acetylcholine at that time and thus activated the muscles. Even today, this neurotransmitter serves to activate the muscles.
So that the uninterrupted signal rotation did not lead to overloading of the neurons involved, an inhibitory back-projection developed from the septum to the amygdala, so that the constant signal rotation was interrupted for the time it took a signal to travel from the amygdala via the hippocampus to the septum. Once this last action potential was used up, the signal rotation started again, because there were more action potentials in the closed loop than the septum could inhibit.
The hippocampus thus became the rotation memory of the nervous system. Once a signal had found its way into the rotation memory, it was available until it was deleted by another algorithm.
For example, all retinal signals were fed into the hippocampus. There, a complete retinal image then rotated in loops, for each image pixel and each visual modality (brightness, colour) there was exactly one closed signal loop.
However, the signals cached in the hippocampus did not directly reach the conscious mind, which evaluated the current present. The past was made available to the subconscious. This was located in the cerebellum.
Therefore, the limbic rotation output also reached the cerebellum via the anterior thalamus during each rotation run. There, the rotation signals are constantly available. And via the cerebellum, they also returned to their places of origin in the cortex. After all, they were - back in early evolutionary times - identical to the original signals.Many millions of years later, however, a special phenomenon emerged.
If you closed your eyes (or alternatively looked at a white sheet of paper), the rotation signals of the limbic system appeared in front of your mind's eye - but this time in the complementary colour. Why is this so?
This is a consequence of the signal divergence in the nucleus olivaris, which developed in the course of evolution. It probably led to changes in the signals.
The output of the retina is sent directly to the limbic system where it participates in signal rotation, at each revolution it also reaches the cerebellum via the anterior thalamus through the nucleus olivaris.
When signal divergence was also established in the nucleus olivaris, the signals in the divergence module of the olive were treated as divergent as in the cortical divergence module. Colour, for example, was encoded as a maximum-encoded neuron vector.
However, a problem arose: the dentate nucleus inverted the cerebellum output. This turned it into a minimum coded signal vector. This signal inversion transferred the current colour into the complementary colour. This reaches - when the eyes are closed or a white sheet of paper is seen - the cortex again via the thalamus and evokes the perceived complementary colour there. The reason for this is the signal inversion in the dentate nucleus and the signal divergence in the olivary nucleus, which did not exist in early prehistoric times. These false colour images are a tribute to the further development of the spinocerebellum into an inversion circuit. Afterimages in complementary colour are an indication of the correctness of my theory.
The rotation signals of the limbic system mainly reach the cerebellum, which represents the subconscious in the brain system. There, the limbic signals, which mostly represent the past, which can also be much longer ago, can be combined with present signals and thus form new complex signals. This is the basis for our long-term memory.
More cannot be presented here because the four-week period during which my wife was at the spa is now almost over. There is only time left for proofreading.However, the last chapter on language acquisition and control is still being completed, but only in short form.
Monograph of Dr. rer. nat. Andreas Heinrich Malczan