Human brain theory
ISBN 978-3-00-068559-0
Monograph of Dr. rer. nat. Andreas Heinrich Malczan
13 The three subsystems of the human brain
Signal processing in the human and vertebrate brain is based (among other things) on three principles, three signal classes and three subsystems.
Principle of the divergence modules
Through divergent distribution of neuronal signals and the action of elementary natural laws, new signal classes arise in the brain. In the case of undamped signal propagation, the phase differences can be detected in a phase module and used to analyse the direction of sound.
In damped signal propagation with non-linear damping, extreme value coded signals can be obtained. If the neuronal transfer function is strictly concave, maximum-coded signals are obtained. This is the preferred mode of operation of the modules in the primary sensory cortex areas of the human brain. The position of the extreme values encodes the values of the primal variables involved, which in turn generate the receptor output. The output of the divergent modules of different modalities forms new submodalities, which form new floors (segments) in the brain.
Principle of transformation and retransformation
If a signal transformation is carried out by a module in the signal path between a receptor and its motor target, a module inverse to this must reconstruct the original signals as well as possible from the new signals by means of a reverse transformation and feed them into the original signal path, while the new signals form a higher-level system in a new evaluation loop.
Principle of feedback neural networks in the pontocerebellum
The pontocerebellum is able to substitute the activity of a cortical neuron population with the activity of a single neuron. A complex signal is thus assigned to a set of active elementary signals.
In the inverse pontocerebellum, the complex signals are transformed back into their active elementary signals.
The pontocerebellum recognises the complex signal belonging to a neuron population even if not all of its elementary signals are active. However, the complex signal then has a lower firing rate, which increases with the number of active elementary signals.
The feedback of the recognised complex signal into the inverse pontocerebellum leads to the activation of all elementary signals assigned to it, even those that were previously inactive. The additionally activated elementary signals represent the self-created input that reaches consciousness in the cortex. The elementary signals activated in this way can activate new complex signals with other elementary signals also created by signal completion. Thus, the brain is able to create its own signals that reach consciousness. These self-generated signals reach both the primary and secondary cortex areas where we become aware of them. In this respect, the pontocerebellum is the material seat of the subconscious mind. This is the basis of our consciousness. It arises in us through the ability to recognise and complete incomplete complex signals.
The constant signal rotation in this feedback system, which is associated with the constant change of signal forms between the elementary form and the complex form, forms the signal-theoretical basis of intelligence.
There are generally three signal classes in the vertebrate and human brain:
· Analogue signals
· Extreme value coded signals
· Complex signals
Analogue signals are characterised by their firing rate, it encodes the value of a quantity that causes action potentials in a receptor. In general, the firing rate can increase with the increase of the magnitude, then we speak of the on-type. In mathematics, this corresponds to a strictly monotonically increasing function. With the Off-type, the firing rate decreases with increasing size, the function is strictly monotonically decreasing. Mixed types also exist.
Extreme value coded signals arise in divergence circuits with non-linear propagation characteristic. Here, the curvature of the transmission characteristic plays a major role. In the spatial propagation in divergence modules of the brain, an upwardly curved transmission characteristic occurs, which is also called strictly concave. When several input sources are superimposed, it leads to a well-defined maximum, which clearly encodes the strength of the primal quantities.
The extreme value coded signals can be transformed back into their output signals in convergence circuits, whereby a certain, usually small signal error also occurs.
Analogue and extreme value coded signals can be used to form complex signals in the pontocerebellum. In the inverse cerebellum, they can be transformed back into the output signals.
Three subsystems in the brain correspond to the three signal classes.
· The primary subbrain processes analogue signals, transmits them to the secondary subsystem and receives the analogue signals from the secondary system, which are extracted from the extreme value coded signals by convergence modules.
· The secondary subbrain receives the analogue signals from the primary subbrain and subjects them to signal divergence, resulting in extreme value coded signals. These are passed on to the tertiary subbrain. On the descending side, the secondary subbrain receives the extreme value coded signals from the inverse pontocerebellum.
· The tertiary subbrain processes elementary signals that belong to analogue or extreme value coded signals after they have been subjected to lateral neighbour inhibition and are thus sparsely coded. It assigns a complex signal to the active neuron groups, this is done in the pontocerebellum. On the descending side, the original elementary signals are recovered from the complex signals and passed on to the secondary system.
Each of these substructures is able to feed its output into the limbic rotation loops so that they are temporarily active there and can be combined with present signals. We refer to this type of memory as rotational memory. Via the nucleus anterior of the thalamus, the limbic signals are available to the system again.
Similarly, each of these three subsystems can feed its output into the basal ganglia, which produce an inhibitory and time-delayed copy of these signals, which is overlaid with present signals to detect signal changes and movements in all modalities. Similarly, an excitatory and time-delayed copy is made. This is done with the substantia nigra pars reticularis as the last station of the basal ganglia. The excitatory and time-delayed copy reaches the pontocerebellum and can be combined with present signals to form new complex signals.
Due to the constant signal passing from the lower to the upper systems on the ascending side and the passing of the signals from top to bottom in the descending side, all three signal classes are available at the same time. Since the output of one system is simultaneously the input of the neighbouring system, a signal rotation occurs.
Complex signals are characterised by the activity of a single neuron. Therefore, they can be fed into the primary system where they are equated to the analogue signals and produce further analysis results as a new input.
Principle of signal rotation
While in the ascending system signals are transformed into the superordinate signal class by transformation in modules, in the descending system the re-transformation of these signals into the original signal class took place, they are fed to the original system as input. This results in constant signal rotation in the system with constant change of signal classes. (Malczan's oscillation theorem, earlier monograph).
During signal oscillation in the system, the incompletely recognised complex
signals are supplemented and completed by activating the missing elementary
signals in the inverse cerebellum. In this way, the system generates its own
input, which rises from the inverse cerebellum and arrives in the cortex, where
we become aware of it.
Principle of supplementing incompletely recognised complex signals
Complex signals are also recognised as such if at least half of the elementary signals belonging to the signal are active. The recognition signal is a complex signal which is transformed into its elementary signals in the descending system. In the process, all components of the elementary signals assigned to it are activated, including the previously inactive ones. In this way, a signal completion constantly takes place in the brain system, in which the non-active signal components of an incomplete but recognised signal are activated, in that the active complex signal in the descending system again activates all elementary signals belonging to it. These elementary signals activated in the descending signal path, which were previously inactive, represent the supplementary signals created by the system itself. They emerge from the subconscious by activating new complex signals in the cerebellum with other elementary signals also induced by signal supplementation, whose components they represent in the elementary form. This is the principle of the emergence of thoughts, feelings, views, ideas and the basic algorithm of all intelligence.
The three-stage brain system of vertebrates and humans is shown below.
I assume that human intelligence is produced precisely by this system.
The most important difference between the brain of humans and the brains of other vertebrates is that they were able to add additional association ladders to the original vertebrate rope ladder system.
The development of the language allowed him to compress the signals.
As an example, compare the storage space needed to store a complete film (where data compression must not be applied) with the storage space needed by the script belonging to the film. Then it quickly becomes clear that data compression through the invention of language leads to the saving of neuronal storage space.
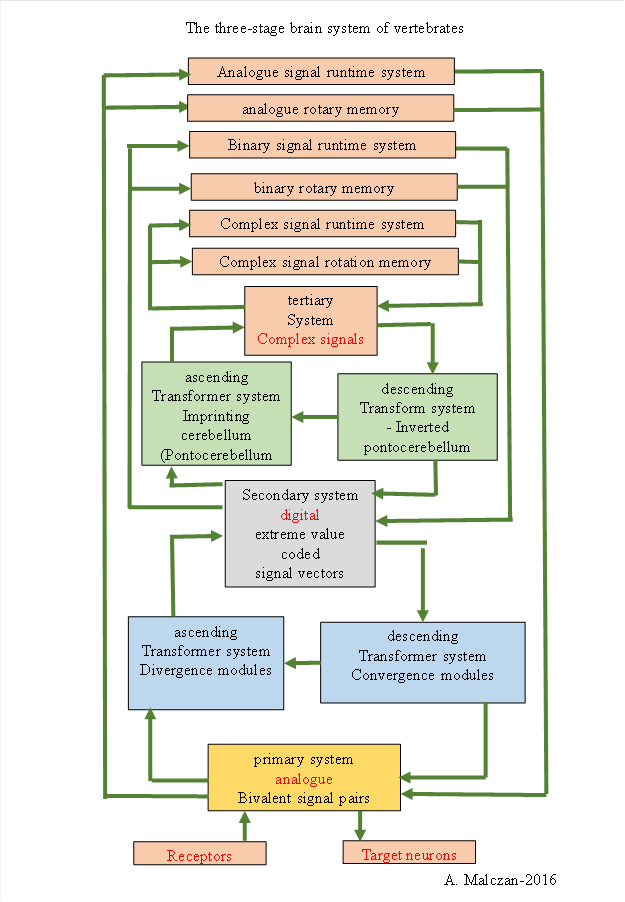
Figure 52: The three subsystems of the brain: primary, secondary and tertiary system