Gehirntheorie des Menschen
ISBN 978-3-00-068559-0
Monografie von Dr. rer. nat. Andreas Heinrich Malczan
14.5 Die Bewegungssteuerung durch Schwerpunktmodule
Die Wirbelsäule lässt Bewegungen in alle Richtungen zu. Der Körper des Menschen kann nach vorne oder nach hinten gebeugt werden, ebenso nach links oder nach rechts. Man kann den Oberkörper auch kreisen lassen.
Eine ähnliche Beweglichkeit kann man bei Fischen beobachten, aber auch bei Schlangen, ebenso auch bei Würmern.
Eine solche Beweglichkeit setzt bei Wirbeltieren Gelenke voraus, die mehrere Freiheitsgrade haben. Modellhaft nehmen wir an, zwei Muskelpaare würden (im Minimalfall) ausreichen, um derartige Bewegungen zu ermöglichen. Ein Muskelpaar ermöglicht die Bewegung in einer Ebene, das zweite Muskelpaar in einer zweiten, dazu geneigten oder dazu senkrechten Ebene.
Vier Muskeln benötigen vier Steuersignale. Jeweils zwei Muskel bilden ein Muskelpaar von antagonistischen Muskeln, die zueinander invers erregt sind. Für sie benötigen wir ein Paar von neuronalen Signalen, die zueinander invers sind.
Für das zweite Muskelpaar benötigen wir ebenfalls ein solches Paar von zueinander inversen Signalen.
Wer kann die passenden Signale liefern?
Zur Beantwortung dieser Frage wenden wir uns dem Zweck der Muskelbewegung zu. Gelenke mit mehr als einem Freiheitsgrad ermöglichen Drehbewegungen. Der Rumpf kann gedreht werden, ebenso der Kopf. Aber auch die Arme und die Beine kann man bei Bedarf im Kreis rotieren lassen. Meist wird eine solche Bewegung von den Augen kontrolliert. Hält man die Bewegung an, so erreicht man eine Beugung in eine gewisse Richtung.
Wir versuchen, die Signale der Augen für die Bewegungssteuerung eines ausgestreckten Arms zu nutzen, der beispielsweise einen Apfel ergreifen soll. Modellhaft untersuchen wir die Bewegung bei einem Menschen, da dessen Gehirnstrukturen bestens erforscht sind.
Die Ausgangssignale entstammen der Netzhaut. Wir entscheiden uns beispielhaft für das rechte Auge. Seine Signale treffen über den Sehnerv im sensorischen Thalamus ein und erreichen den primären visuellen Cortex, aber auch eine Struktur, die als Tectum opticum bekannt ist. Bei Wirbeltieren wird diese auch als Colliculus superior bezeichnet und ist bilateral vorhanden. Dort treffen jedoch (vermutlich) nur die Signale der magnocellularen Netzhaut-Ganglienzellen mit besonders großen rezeptiven Feldern ein. Insofern wird im Tectum opticum eine Mittelwertabbildung der Netzhaut vorliegen.
Die Axone der visuellen Projektionsneuronen in das Tectum mögen in einer Fläche enden, die ein Mittelwertabbild der Netzhaut darstellt. Bei höheren Wirbeltieren kann es - bedingt durch die Ausbildung unterschiedlicher visueller Rezeptorarten - auch durchaus mehrere solcher Eingangsflächen geben, etwa eine für Hell/Dunkel, möglicherweise sogar für verschiedene Farben. Dann sind diese in Schichten übereinander gestapelt. Wir entscheiden uns für die tectale Inputfläche Dunkel-On/Hell-Off. Sie reagiert auf dunkle Objekte vor hellem Hintergrund.
Dort enden die Axone der visuellen Mittelwertneuronen dieser Sehmodalität in einer nahezu kreisförmigen Fläche, die wir uns als Mittelwertabbild der Netzhaut vorstellen können. Wir unterstellen, dass bei der Übertragung der visuellen Signale in das Tectum opticum die Abbildungstreue erhalten bleibt, die Abbildung also retinotop ist.
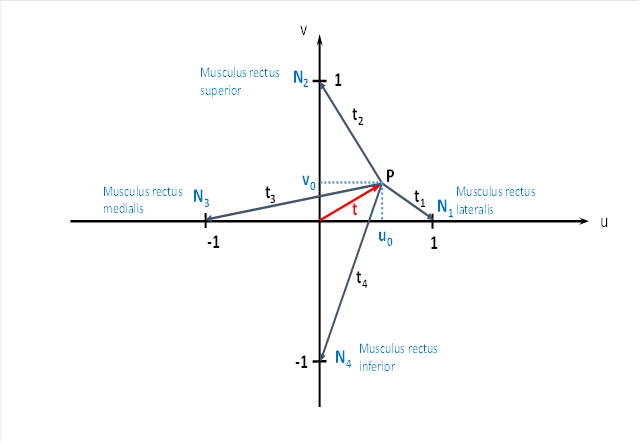
Wir denken uns in im Tectum opticum ein virtuelles Koordinatensystem aus einer u-Achse und einer v-Achse. Die u-Achse verlauft waagerecht durch die Sehfeldmitte, die v-Achse senkrecht ebenfalls durch die Sehfeldmitte. Blicken wir genau geradeaus, so können wir dem Blickpunkt die Koordinaten u = 0 und v = 0 zuordnen. Wir entscheiden uns für die Variablen u und v, weil wir später ein zweites Koordinatensystem mit einer x-Achse und einer y-Achse benötigen werden.
Denken wir uns in der Sehfeldebene einen Kreis mit einem gewissen Radius, dessen Mittelpunkt durch die Sehfeldmitte verläuft, so wird dieser Kreis über die visuelle Projektion von der Retina zum Tectum opticum dort ebenfalls als Kreis in der Projektionsfläche abgebildet.
Nun möge der visuelle Hintergrund eine weiße Wand sein, auf der ein kleiner schwarzer Kreis (beispielsweise ein kleiner schwarzer Ball) genau auf einer Kreislinie um das Sehfeldzentrum rotiert. Dies könnte man beispielsweise dadurch erreichen, dass man dieses Geschehen im Computer visuell simuliert und über einen Projektor an die Wand projiziert.
Das Bild des rotierenden schwarzen Balls im Tectum kann nun durch zwei Größen beschrieben werden. Er befindet sich im Punkt P. Erstens hat er einen Abstand t vom Zentrum. Zweitens hat die Verbindungsgerade von P zum Zentrum einen Winkel ω mit der u-Achse. Alternatix kann man auch einfach die u-Koordinate und die v-Koordinate des Apfelmittelpunktes angeben. Beide hängen mit dem Abstand und dem Winkel über den Satz des Pythagoras und über trigonometrische Funktionen zusammen.
Hierbei gilt
![]()
![]()
Wir bezeichnen den Winkel ω als Phasenwinkel und t als Radiusvektor im Urgrößendiagramm.
Wir erinnern uns daran, dass das Tectum eine geschichtete Struktur darstellt, in den oberen Schichten endet der Retinaoutput getrennt nach Modalitäten: Hell-On/Dunkel-Off, Dunkel-On/Hell-Off, Rot-On/Grün-Off, Grün-On-Rot-Off, Blau-On/Gelb-Off.
Wir konzentrieren uns auf die waagerechte und kreisrunde Schicht der retinalen Signale Dunkel-On/Hell-Off, die auf schwarze Objekte vor weißem Hintergrund am besten reagiert. Hätte der Hintergrund eine andere Farbe und der Ball eine andere, komplementäre Farbe dazu, wären die mathematischen Überlegungen identisch, würden sich aber auf eine andere visuelle Inputschicht des Tectums beziehen.
Wir gehen also davon aus, dess der schwarze Ball an der Stelle der Retina, wo sich sein Abbild befindet, die Rezeptoren maximal erregt. Alle übrigen Rezeptoren des Typs Dunkel-On/Hell-Off sind völlig inaktiv.
Eine retinale Ganglienzelle in der Mitte des Abbildes möge mit ihrem größeren Dendritenbaum die Retinaerregung des Auges aufgreifen und zum Tectum senden. Die übrigen retinalen Ganglienzellen seine inaktiv.
Daher tritt an der Stelle P des Tectums die visuelle Erregung ein, alle übrigen Inputneuronen in dieser Tectumschicht erhalten kein Signal. (Wir betrachten hier das einäugige Sehen und daher auch nur ein Tectum von zweien).
Nun wird sich die Erregung vom Punkt P über Interneuronen zu den vier Neuronen ausbreiten, die wir in der Abbildung mit N1, N2, N3 und N4 bezeichnet haben. Jedes Neuron erhält einen Erregungsanteil, der von der Entfernung zum Punkt P abhängt. Diese vier Entfernungen haben wir mit t1, t2, t3 und t4 bezeichnet.
Warum sind diese Neuronen an den Stellen, wo wir sie eingezeichnet haben? Die eingezeichneten Neuronen senden Ihre Signale zu vier Augenmuskeln sowie zu vier Halsmuskelgruppen. Weitere Projektionsziele werden wir später benennen.
Wir beziehen uns hier auf das rechte Auge.
Das Neuron N1 projiziert zum Augenmuskel Musculus rectus lateralis, der den Blick nach außen wendet. Gleichzeitig kann dieses Neuron die Halsmuskelgruppe aktivieren, die den Kopf nach rechts dreht.
Das Neuron N2 sendet sein Axon vom Tectum opticum aufwärts zum Augenmuskel mit dem Namen Musculus rectus superior. Dieser bewirkt bei Kontraktion eine Bewegung des Auges nach oben, der Blick wandert also in Richtung der u-Achse nach oben. Eine kleine Auslenkung des Augapfels nach innen wollen wir hier vernachlässigen, da es nur um das Prinzip geht.
Nach unten projiziert das Neuron N2 zu denjenigen Halsmuskeln, die eine Hebung des Kopfes nach oben bewirken.
Das Neuron N3 aktiviert den Augenmuskel Musculus rectus medialis, der den Blick nach innen wendet (wenn wir das rechte Auge als Grundlage nehmen). Ebenso aktiviert dieses Neuron diejenigen Halsmuskel, die eine Kopfdrehung nach links bewirken.
Das Neuron N4 des Tectums erregt einerseits beim Auge den Musculus rectus inferior, der eine Augenbewegung nach unten bewirkt, so dass der Blick gesenkt wird. Gleichzeitig werden die Halsmuskel erregt, die den Kopf senken.
Hätten wir uns für das linke Auge entschieden, so wären links und rechts zu vertauschen!
Doch woher nehmen die Augenmuskel ihre Erregungen her? Es ist ganz einfach. Die Erregungen der Augenmuskel werden vom Auge verursacht. Die visuellen Mittelwertsignale der Netzhaut projizieren in die visuelle Eingangsschicht des Tectums und erregen dort die visuellen Eingangsneuronen, die in dieser Fläche ebenso verteilt sind wie die Mittelwertneuronen des Augen. So bleibt die Abbildungstreue erhalten, die Abbildung ist retinotop.
Über Interneuronen breitet sich die Erregung vom Erregungszentrum P zu den vier Outputneuronen des Tectums aus. Hierbei tritt während der Ausbreitung der Erregung eine Signaldämpfung auf, die exponentiell mit dem Quadrat der Entfernung zunimmt. Wir wollen die Erregung der vier Outputneuronen berechnen.
Als Inputerregung nehmen wir eine Feuerrate fp, die von der Retina kommend im Punkt P des Tectums eintrifft und stellvertretend das Bild des schwarzen Balls repräsentieren möge. Vom Punkt p breitet sich die Erregung fp zu den vier Outputneuronen aus.
Da die Erregung, die die Neuronen N1, N2, N3 und N4 erreicht, vom Quadrat des Abstandes zum Punkt P abhängt, berechnen wir zunächst die Quadrate.
![]()
![]()
![]()
![]()
Bezeichnen wir mit fp die Feuerrate, die im Punkt P von der Retina kommend im Tectum opticum eintrifft, so können wir die Feuerraten f1, f2, f3 und f4 berechnen, die in den Neuronen N1, N2, N3 und N4 ankommen wird.
Die Formeln dazu lauten:
![]()
![]()
![]()
![]()
Den gemeinsamen Faktor
![]()
Klammern wir aus und erhalten
![]()
![]()
![]()
Wir beachten, dass die Größe fm lediglich die Variablen u0 und v0 enthält beziehungsweise den Vektor t , die Größen x oder y kommen hier nicht vor. Die Größe λ ist die Längskonstante der Neuronen.
Leicht ersichtlich ist, dass alle vier Feuerraten periodische Funktionen des Winkels ω sind und die Periode 2π aufweisen.
Rotiert also der schwarze Ball vor weißem Hintergrund um das Sehfeldzentrum, so folgt der Blick diesem Objekt und die Blickrichtung rotiert ebenfalls um das Zentrum, da die Augenmuskelsignale periodische Funktionen sind.
Damit haben wir die Feuerraten der Neuronen N1 bis N4 berechnet, die auf den Axonen dieser Neuronen kopfwärts die zugeordneten Augenmuskeln erreichen und die ebenfalls zu den vier Halsmuskelgruppen für die Kopfbewegung projizieren.
Wir unterstellen, dass sich diese neuronalen Erregungen ohne Veränderung der Feuerrate vom Tectum opticum nach oben und unten auf den Axonen ausbreiten und in verschiedenen Zielneuronen eintreffen.
Solange sich das visuelle Objekt außerhalb der Blickfeldmitte befindet, sind die vom Schwerpunktmodul erzeugten Signale für die Motoneuronen der Augenmuskeln unterschiedlich, so dass eine Bewegung des Auges stattfindet. Erst wenn das Objekt die Bildmitte erreicht, endet die Augenbewegung. Dann besitzen die Signale für die Motoneuronen der Augenmuskeln alle die gleiche Feuerrate.
Wenn also eine schwarze Fliege auf einer weißen Wand hin und her läuft, so steuert sie nicht mit imaginären Kräften die Augenmuskeln. Es ist das visuelle Abbild der Fliege auf der Netzhaut, das die lichtempfindlichen Rezeptoren erregt. Diese Erregung gelangt ins Tectum opticum und breitet sich zu dessen Rand aus, wo sich die Projektionsneuronen zu den Motoneuronen der Augenmuskeln befinden. Durch die unterschiedlichen Abstände des Fliegenabbildes zum Tectumrand ist die Signaldämpfung dieser Signale unterschiedlich. Deshalb erhalten die Augenmuskeln unterschiedliche Kontraktionsbefehle, solange sich die Fliege nicht zentral im Sehfeld befindet.
Auf ähnliche Weise steuert ein vestibuläres Schwerpunktmodul im Torus semicircularis die Muskeln des Körpers, damit wir möglichst in einer stabilen Lage verharren und nicht hinfallen. Hier ist die Steuerung komplexer, da viele Gelenke in Reihe geschaltet sind und Gelenkwinkel gegeneinander verrechnet werden müssen. Dazu sind geschichtete Schwerpunktmodule durchaus in der Lage. Der mathematische Beweis ist jedoch etwas komplizierter und wird hier nicht gebracht.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan