Gehirntheorie des Menschen
ISBN 978-3-00-068559-0
Monografie von Dr. rer. nat. Andreas Heinrich Malczan
14.7 Das Modul für stereoskopisches Sehen
Wirbeltiere besitzen zwei Augen. Dadurch sind sie in der Lage, räumlich zu sehen. Deshalb müssen sie in ihrem Gehirn ein Modul besitzen, welches räumliches Sehen ermöglicht. Vielleicht sind auch mehrere Module dafür nötig. Denn am Sehprozess sind mehrere Subsysteme beteiligt. Einerseits steuern Augenmuskel die Bewegung der Augen. Weiterhin muss es eine Steuerung für die Pupullen geben. Und zu guter Letzt muss die Form der Augenlinse je nach Entfernung des anvisierten Objekts so verändert werden, dass dieses Objekt auf der Netzhaut in bestmöglicher Schärfe abgebildet wird. Aber auch die Pupillen müssen angesteuert werden, um die Augen an die bestehende mittlere Helligkeit anzupassen.
Doch das visuelle System hat weitere Funktionen. So ist es zum Beispiel für Raubtiere erforderlich, die Entfernung zum Objekt zu ermitteln, damit man abschätzen kann, ob es als Beute in Frage kommt oder wegen seiner großen Fluchtgeschwindigkeit nicht mehr erreicht werden kann.
Ebenso wichtig ist die Richtungsbestimmung zum Objekt. Noch komplizierter wird diese bei denjenigen Raubvögeln, deren Beutetiere ebenfalls fliegende Vögel sind.
Und letztlich bilden die visuellen Signale die Grundlage für den Orientierungssinn. Welche Algorithmen mögen der Tatsache zugrunde liegen, dass ein Hamster nach der Futtersuche auf dem Feld seinen eigenen Bau wiederfindet?
Es muss im Gehirn der Wirbeltiere und des Menschen Subsysteme geben, die all diese Aufgaben bewältigen können. Der Autor wird zeigen, dass die Grundlage für all diese Fähigkeiten von gewöhnlichen Divergenzmodulen gelegt werden, wobei deren Output sowohl im Cerebellum, in den Basalganglien und letztlich auch im Cortex weiterverarbeitet wird.
Wir werden nicht alle visuell wirksamen Module beschreiben können. Allerdings muss darauf hingewiesen werden, dass diese Beschreibungen zunächst hypothetisch sind. Der Nachweis, dass diese real existieren, müssen andere erbringen. Hier wird lediglich aufgezeigt, welche Algorithmen die Natur anwenden könnte, um solche Ziele erreichen zu können.
Ausgehend vom frühen Strickleiternervensystem unterteilen wir die neuronalen Kerne in sensorische Kerne und motorische Kerne, diese befinden sich jeweils auf beiden Körperhälften des Lebewesens. Zusätzlich gibt es sensorische und motorische Seitenwechselkerne.
Die visuell wirksamen Kerne, die vorwiegend der Steuerung der Augenmuskel dienen, befinden sich im segmentierten Strickleitersystem bzw. seinem Nachfolger mit Sicherheit in der tectalen Etage.
Im Kapitel "Die Bewegungssteuerung durch Schwerpunktmodule" wurde bereits gezeigt, wie das sensorische Tectum opticum als Schwerpunktmodul arbeitet und die Signale für die vier geraden Augenmuskeln erzeugt, mit denen der Blick sowohl seitlich als auch in der Höhe gesteuert werden kann. Ebenso eignen sich diese tectalen Outputsignale zur Steuerung der Halsmuskeln, teils auch zur Steuerung der Rumpfmuskeln (Rumpfdrehung zur visuellen Objektverfolgung) oder sogar zur Steuerung der Armbewegungen, um etwa ein Objekt zu ergreifen.
Wie aber kann man das räumliche Sehen erklären und die Fähigkeit, sich ein räumliches Abbild von der Umgebung zu machen, bei dem die erkannten Objekte hinsichtlich ihrer Lage und ihres Abstandes in einer Art Karte abgespeichert werden?
Eine solche Karte ist ein aktuelles Abbild der Umgebung. Sobald das Lebewesen seinen Ort verändert, ist die bisherige Karte ungültig und eine neue entsteht.
Es gibt aber auch eine neuronale Karte, die bei Ortsveränderungen des Lebewesens erhalten bleibt, weil sich der Standort des Lebewesens in der Karte ändert, sobald es sich bewegt.
Und letztlich gibt es auch eine neuronale Karte, welche die Umgebung abbildet, aber so, dass die Karte sich dreht, sobald das Lebewesen sich dreht. So wie es eine Karte in einem GPS-gesteuerten Navigationsgerät auch tut. So weit werden wir in dieser Monografie wohl nicht kommen, da die dafür erforderlichen neuronalen Algorithmen vom Autor noch nicht vollends verstanden wurden.
Wir werden hier - wahrscheinlich erstmalig - ein visuelles Modul für räumliches Sehen entwerfen und zeigen, wo es sich im Gehirn befindet. Hierbei sollte die Gehirnarchitektur möglichst realitätsnah berücksichtigt werden. Der große Mangel vieler Gehirntheorien besteht darin, den realen Aufbau des Gehirns und des Nervensystems zu vernachlässigen und rein theoretisch an das Problem heranzugehen.
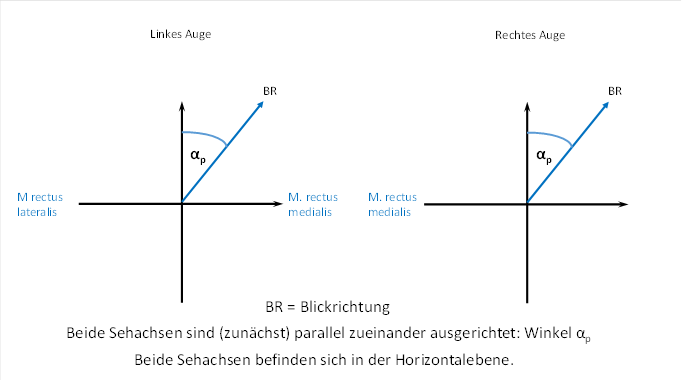
Wir diskutieren zunächst den Fall, dass sich die Sehachse beider Augen in einer waagerechten Ebene befindet, während der Kopf die Normallage einnimmt. Wir schauen also waagerecht nach vorn oder etwas zur Seite, ohne den Blick zu heben oder zu senken.
Zunächst schauen wir genau geradeaus in die Ferne, quasi ins Unendliche. Dabei werden beide Sehachsen beider Augen parallel zueinander ausgerichtet.
Nun wenden wir den Blick etwas nach rechts und schauen ebenfalls in die Ferne, so dass auch diesmal beide Sehachsen parallel zueinander ausgerichtet sind.
Hierbei tritt der Winkel αp auf. Er soll der Winkel sein, den die Sehachse mit der Blickrichtung bildet, wenn wir geradeaus schauen. Dies ist in der folgenden Abbildung zu sehen. Wir nennen den Winkel αp, weil er sich auf die Sicht in die Ferne bezieht, bei der die Sehachsen beider Augen parallel zueinander ausgerichtet sind.
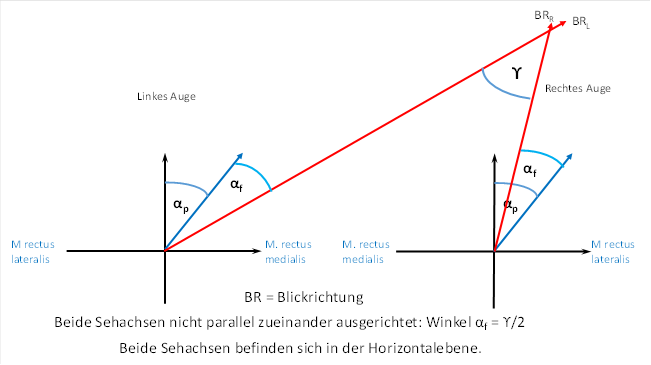
Nun entschließen wir uns, mit unseren Augen ein Objekt zu fixieren, welches sich auf der rechten Seite befindet und nur wenige Meter von uns entfernt ist. Hierbei sind die zwei Sehachsen nicht mehr paralel zueinander. Für das linke Auge müssen wir den Winkel αp um einen zusätzlichen Winkel αf vergrößern. Wir bezeichnen den zusätzlichen Winkel mit αf, weil er beim Fokussieren auf ein Objekt in der Nähe nötig ist. Für das rechte Auge müssen wir den Winkel αp um einen den Winkel αf verkleinern. Hierbei zeigt sich, dass der Winkel αf genau die Hälfte des Winkels ϒ beträgt. Der Winkel ϒ ist der Schnittwinkel beider Sehachsen.
Dies zeigt die nachfolgende Abbildung.
Wir bezeichnen den Auslenkwinkel von der Geradeaus-Stellung für das linke Auge mit αL und für das rechte Auge mit αR. Dies ist in der folgenden Abbildung dargestellt.
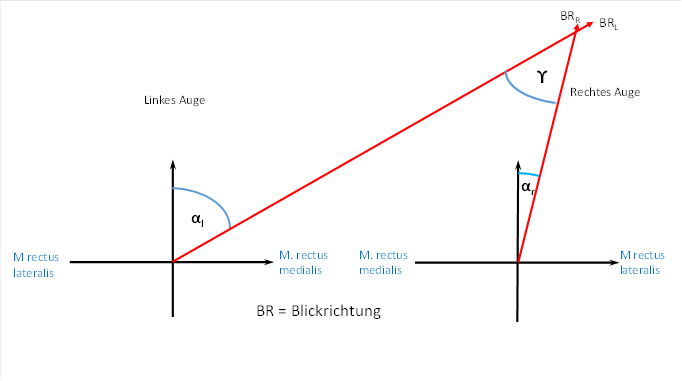
Ein Vergleich der beiden Darstellungen ergibt die gesuchten Formeln für die Blickrichtung.
![]()
![]()
![]()
Wir brauchen nun einen theoretischen Ansatz für die Feuerrate der beteiligten Augenmuskeln. Wir gehen von einer mittleren Feuerrate fm aus. Diese soll vorhanden sein, wenn die Augen genau geradeaus schauen, ohne den Blick zu heben oder zu senken.
Schauen wir nach rechts in die Blickrichtung, die durch den Winkel α vorgegeben wird, so muss derjenige Augenmuskel stärker kontrahieren, der den Blick nach rechts lenkt. Für das linke Auge ist dies der Musculus rectus medialis, für das rechte Auge dagegen der Musculus rectus lateralis. Der jeweilige motorische Gegenspieler muss dagegen seine Kontraktion etwas verringern.
Konkrete Formeln für die Kontraktion der Augenmuskeln fehlen in den bisherigen Theorien. Wir gehen davon aus, dass die Feuerrate exponentiell mit dem Auslenkwinkel wächst, wobei wir die Längskonstante λ der Neuronen in den Formeln unterbringen.
Wir verwenden den Index L für den Musculus rectus lateralis, den Index M für den Musculus rectus medialis.
Dann möge folgender Ansatz für die Feuerraten f verwendet werden:
|
Musculus rectus |
Feuerrate |
|
|
links |
medialis |
|
|
links |
lateralis |
|
|
rechts |
lateralis |
|
|
rechts |
medialis |
|
Derjenige Muskel, der mehr angespannt ist, hat im Exponenten das Plus-Zeichen. Der weniger angespannte Muskel ist am Minuszeichen im Exponenten zu erkennen.
Wir erkennen, dass bei beiden Augen die Feuerrate des Musculus rectus lateralis invers zur Feuerrate des Musculus rectus medialis ist. Beide sind zueinander als motorische Gegenspieler einzuordnen.
Aus diesen vier Feuerratensignalen wollen wir ein Modul konstruieren, welches räumliches Sehen ermöglicht und welches in einer realen Struktur des Gehirns tatsächlich vorkommt.
Doch zuvor müssen wir uns an die zwei weiteren Arten von Augenmuskeln erinnern, da auch ihre Signale zu berücksichtigen sind. Es ist der Musculus rectus superior und der Musculus rectus inferior. Der erste hebt den Blick, der zweite senkt ihn. Wir nehmen an, dass der Blick um den Winkel β angehoben wird. Dieser Winkel ist identisch für das linke und das rechte Auge.
Dann möge der folgende Ansatz den Zusammenhang zwischen dem Winkel und der Feuerrate bestehen:
|
Musculus rectus |
Auge links |
Auge rechts
|
Wirkung |
|
superior |
|
|
Blick heben |
|
inferior |
|
|
Blick senken |
Damit haben wir 8 Augenmuskelsignale, vier vom rechten Auge und vier vom linken Auge. Nun müssen wir nur noch klären, auf welche Art diese Signale in einem Modul oder einem neuronalen Kern angeordnet werden könnten.
Da es sich um Signale beider Körperseiten handelt, gibt es nur eine Art von Kern, der beide Signalversionen verarbeitet. Dieser Kern muss ein Seitenwechselkern sein.
Bereits beim urtümlichen Strickleiternervensystem gab es in jeder Etage und auf jeder Körperseite je einen motorischen und einen sensorischen Seitenwechselkern. Über sie zogen die entsprechenden Signale einer Körperseite zur Gegenseite. Damit war der Signalaustausch zwischen beiden Körperseiten möglich.
Die Signale der ipsilateralen Körperseite zogen zum Seitenwechselkern, um auf die Gegenseite zu gelangen. Ebenso zogen die Signale der kontraleteralen Seite zum Seitenwechselkern, um auf die ipsilaterale Seite zu kommen. Dadurch gibt es hier eine Begegnung dieser Signale, so dass neuronale Strukturen auf beide Körperseiten zugreifen konnten.
Es gibt sowohl sensorische als auch motorische Seitenwechselkerne. Wir untersuchen hier die motorische Seitenwechselkerne für die Signale der Sehnenorgane der geraden Augenmuskeln beider Augen.
Die Signale des linken Auges ziehen zum linken Seitewechselkern, die des rechten Auges zum rechten Seitenwechselkern. Beide befinden sich auf der motorischen Seite des alten Strickleiternervensystems bzw. des Neuralrohrs in der Etage, in der sich das Tectum opticum befindet, welches die Augenmuskelsignale auswertet.
Stellt man sich dies im Geist räumlich als Modell vor, so bemerkt man die enge Nachbarschaft beider Kerne.
Die Natur hat es sicherlich so eingerichtet, dass die zwei verschiedenen motorischen Seitenwechselkerne der beiden Körperseiten so eng benachbart sind, dass sie letzlich im Verlauf der Evolution verschmelzen konnten. Dann gibt es nur noch einen gemeinsamen motorischen Seitenwechselkern in jedem Segment des segmentierten Nervensystems, der gleichzeitig Input von der linken und von der rechten Körperseite erhält.
Wir können uns diesen gemeinsamen motorischen Seitenwechselkern - der mit Sicherheit in der tectalen Etage angeordnet ist - als einen Würfel vorstellen. Die vier Signale des linken Auges enden an der linken Würfelfläche in den vier Ecken. Die Signale des rechten Auges enden entsprechend in den vier rechten Ecken des Würfels.
Wir drehen den Würfel für die weitere Analyse derart, dass die Signale für die waagerechte Augenbewegung ebenfalls in der waagerechten Ebene liegen. Dies erleichtert die späteren Betrachtungen.
Weiterhin drehen wir den Würfel so, dass die zwei Ecken, welche die Signale der Muskeln empfangen, die den Blick heben, sich oben befinden. Die zwei anderen Ecken empfangen die Signale der Muskeln, die den Blick senken und befinden sich unten.
Damit hat der Würfel die Lage, die wir für unsere Analyse benötigen.
In das Zentrum des Würfeld legen wir den Ursprung eines räumlichen Koordinatensystems. Die x-Achse verlaufe von links nach rechts in der waagerechten Ebene, die y-Achse von vorn nach hinten und die z-Achse von unten nach oben. In den acht Ecken des Würfels sollen die neuronelen Signale der acht Augenmuskeln mit ihren Feuerraten f1 bis f8 eintreffen.
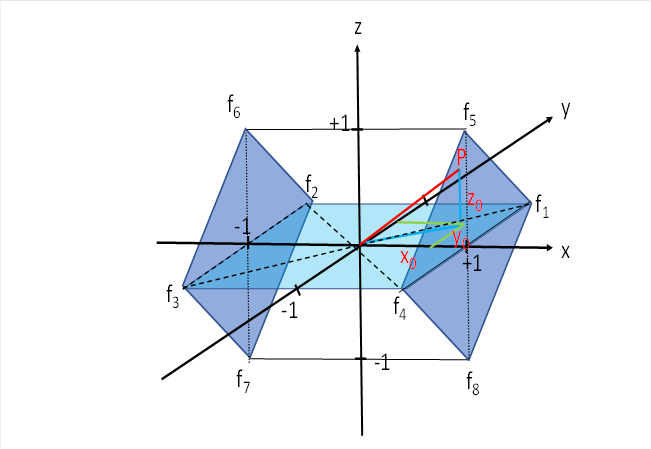
Wir untersuchen zunächst nur vier der acht Signale, indem wir uns auf die Augenmuskeln beschränken, die eine Augenbewegung nach rechts und nach links bewirken. Die Muskeln, die ein Heben oder ein Senken des Blicks bewirken. lassen vor zunächst unbeachtet, wir werden sie erst später in unsere Betrachtung einbeziehen. Damit beschränken wir uns zunächst auf das Quadrat, welches in der x-y-Ebene liegt. Es ist in der obigen Abbildung hellblau.
Der Input der ausgewählten vier Augenmuskelsignale für die seitlichen Augenbewegungen sollen in den vier Punkten P1(1;1), P2(-1;1), P3(-1;-1) und P4(1;-1) eintreffen. Dies ist in der nachfolgenden Abbildung dargestellt. Sie zeigt die x-y-Ebene, auf die wir uns beziehen.
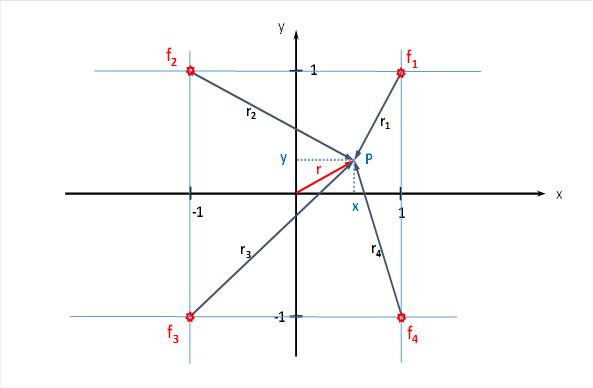
Für die Gesamterregung im Punkt P(x,y) gilt dann die Feuerrate
![]()
Diese vier neuronalen Erregungen breiten sich in der x-y-Ebene nach allen Seiten aus, dabei wird die Erregungsübertragung durch die zahllosen Interneuronen in dieser Struktur realisiert. Damit liegt hier ein Divergenzmodul vor. Hierbei tritt - wie in einem Divergenzmodul üblich - eine exponentielle Signaldämpfung auf, die mit dem Abstand quadratisch zunimmt. Mit der Längskonstanten λ der beteiligten Interneuronen können wir die im Punkt P(x,y) eintreffende Erregung berechnen.
Hat ein Punkt P zu einem Inputneuron mit der Feuerrate f0 den Abstand r, so wird der Zusammenhang zwischen der Inputerregung f0 und der Erregung im Punkt P durch folgende Formel beschrieben:
![]()
Hierbei ist λ die Längskonstante der Interneuronen.
Im Exponenten tritt der Abstand quadratisch auf, weil sich die Erregung nicht mehr auf einem einzelnen Axon ausbreitet, sondern über stark verzweigte Axonbüschel. Dadurch nimmt die Erregung mit dem Abstand schneller ab als auf einem geraden, unverzweigten Axon.
Diese in den Würfelecken eintreffende Erregung wird von einem Neuron im Punkt P (oder in dessen unmittelbarer Nachbarschaft) aufgenommen und stellt den Output des Divergenzmoduls im Punkt P(x,y) dar.
Da die einzelnen Teilsummanden der Gleichung
![]()
im betrachteten Quadrat streng konkave Funktionen in den Variablen x und y darstellen (Stichwort Hessematrix), existiert in dieser Fläche ein globales Erregungsmaximum, welches Neurologen möglicherweise messtechnisch nachweisen könnten. Bei Veränderungen der Augenwinkelstellung würde dieses Erregungsmaximum umherwandern. Uns interessiert, welche Augenwinkel einem Erregungsmaximum zugeordnet werden können. Ist es möglich, aus den Koordinaten x und y des Erregungsmaximums die Winkel αp und αf der Augen zu ermitteln.
Hier müssen wir die Differentialrechnung anwenden, indem wir die partiellen Ableitungen von f(x,y) nach x und y berechnen und gleich null setzen.
Wir berechnen zunächst die Quadrate der Radiusvektoren.
![]()
![]()
![]()
![]()
Wir setzen die Quadrate der Radiusvektoren in die Ausgangsgleichung ein, ziehen jedoch den gemeinsamen Faktor vor die Summe.
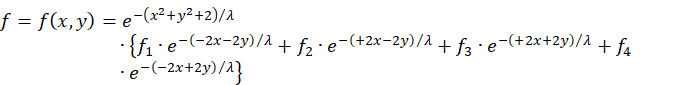
![]()
Nun setzen wir die Feuerraten f1 bis f4 ein.
Wir müssen nur noch festlegen, welches der vier Signale in welchem Punkt eintreffen soll. Wir entscheiden uns dafür, die zwei Signale derjenigen Augenmuskeln, die den Blick nach rechts bewirken, oben anzuordnen. Die Signale, die eine Augenbewegung nach links verursachen, ordnen wir unten ein.
Dabei sind die Signale des linken Auges generell links, die des rechten rechts anzuordnen.
Dann gilt:
![]() rechtes Auge, muscilus rectus
lateralis, Blick nach rechts
rechtes Auge, muscilus rectus
lateralis, Blick nach rechts
![]() linkes Auge, musculus rectus medialis,
Blick nach rechts
linkes Auge, musculus rectus medialis,
Blick nach rechts
![]() rechtes Auge, musculus rectus medialis,
Blick nach links
rechtes Auge, musculus rectus medialis,
Blick nach links
![]() linkes Auge, musculus rectus lateralis,
Blick nach links
linkes Auge, musculus rectus lateralis,
Blick nach links
Der Grund für diese spezielle Anordnung liegt in der Vermutung begründet, es gäbe einen Körperseitenmarker. Daher ordnen sich die Inputneuronen für eine Augenbewegung nach rechts auf der rechten Seite des Quadrats an. Analog befinden sich die Inputneuronen für eine Augenbewegung nach links auf der linken Seite an. Der Körperseitenmarker wird von den betreffenden Muskeln über die Sehnenorgane an die Zielneuronen in dieser tectalen Struktur übertragen.
Das Einsetzen der Feuerraten liefert:
![]()
![]()
Zusammenfassen liefert ein Zwischenergebnis mit Hyperbelfunktionen gemäß
![]() bzw.
bzw. ![]()
Damit ergibt sich:
![]()
Wir wenden folgendes Additionstheorem für Hyperbelfunktionen an:
![]()
Die Anwendung dieses Additionstheorens ergibt:
![]()
![]()
Wegen
![]()
![]()
und
![]()
![]()
ergibt sich die endgültige Formel:
![]()
Diese Formel beschreibt die Gesamterregung im Punkt P(x,y) des tectalen motorischen Seitenwechselkerns bei Beschränkung der Erregung auf die x-y-Ebene.
Wegen der strengen Konkavität dieser Funktion im betrachteten Quadrat mit der Seitenlänge 2 existiert dort ein globales Maximum.
Durch Nullsetzen der partiellen Ableitungen nach x und nach y kann man Bedingungen für das globale Maximum herleiten.
Wir berechnen die partielle Ableitung nach x.
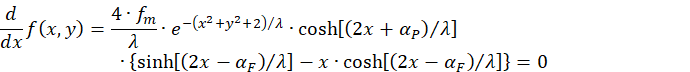
Wegen
![]()
muss folgende Bedingung erfüllt sein, damit die partielle Ableitung nach x gleich null wird:
![]()
Dies ist gleichbedeutend mit
![]()
Wir formen um:
![]()
![]()
Damit haben wir die ersten Bedingung für das Vorliegen eines Maximums der Erregungsfunktion f = f(x,y) im Punkt p(x,y) erhalten, sie lautet:
![]()
Eine zweite Bedingung ergibt sich, wenn wir die partielle Ableitung der Funktion nach y gleich null setzen:
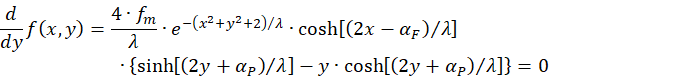
Auch hier gilt:
![]()
Somit verbleibt als Bedingung für ein Maximum nur noch die Gleichung
![]()
Umformung ergibt die zweite Bedingung für das Vorliehen eines Maximums:
![]()
Endergebnis:
Die Erregungsfunktion
![]()
des tectalen motorischen Seitenwechselkerns in der x-y-Ebene nimmt im Punkt P(x,y) dann und nur dann ein globales Maximum an, wenn die beteiligten Augenwinkel αF und αP den folgenden Bedingungen genügen:
![]()
![]() .
.
Merke:
In der x-y-Ebene des tectalen
Seitenwechselkerns verschlüsselt der x-Wert des globalen Maximums den
Fokussierwinkel ![]() und der y-Wert den Blickrichtungswinkel
und der y-Wert den Blickrichtungswinkel ![]() beim Blick ins Unendliche.
beim Blick ins Unendliche.
Da der Fokussierwinkel direkt an den Abstand des anvisierten visuellen Objekts gekoppelt ist, verschlüsselt die x-Koordinate des globalen Erregungsmaximums den Objektabstand des anvisierten Objekts.
Doch welchen Nutzen zieht das Lebewesen daraus, dass in der tectalen x-y-Ebene des Seitenwechselkerns in x-Richtung die Objektentfernung und in y-Richtung der (mittlere) Blickwinkel (beim Blick ins Unendliche) maximumcodiert verschlüsselt werden?
Ein solcher Aufwand an Ressourcen muss Vorteile gebracht haben, sonst hätte sich ein solches System nicht herausgebildet. Wobei die Herausbildung recht simpel erfolgte. Die Anzahl der Interneuronen musste zunehmen, gleichzeitig mussten zusätzliche Outputneuronen die übertragene Erregung empfangen. Und die zusätzlichen Interneuronen und Outputneuronen mussten sich möglichst gleichmäßig im Modul verteilen. Den Rest bewerkstelligte die neuronale Signaldämpfung.
Wir wiederholen:
Auch wenn wir uns auf die Betrachtung der x-y-Ebene beschränken, liegt hier ein Divergenzmodul vor. Neben den vier Inputneuronen an den Ecken des Quadrates in der x-y-Ebene gibt es innerhalb dieser Fläche unzählige Projektionsneuronen, die in der Fläche (und im Raum) verteilt sind. Dazwischen befinden sich unzählige Interneuronen in diesem Quadrat. Sie übertragen die Erregung von den vier Inputneuronen zu den hunderten oder gar tausenden Projektionsneuronen. Offenbar findet auch eine Erregungsübertragung von Interneuron zu Interneuron statt, die diese recht klein sind im Vergleich zur Gesamtausdehnung.
Hierbei tritt eine abstandsabhängige Dämpfung auf, die mit dem Abstand zwischen Erreger und Erregtem quadratisch zunimmt. Diese Übertragungsfunktion ist daher innerhalb des Quadrates und in dessen Umgebung streng konkav, also stets als Funktionsfläche nach oben gekrümmt. Daher gibt es in der Fläche genau ein Erregungsmaximum. Dies ist äußerst wichtig!
Bevor der Output diese Struktur verlässt, muss er (höchstwahrscheinlich) durch eine Schicht aus hemmenden Interneuronen. Diese sorgen für eine laterale Hemmung zur Kontrastverstärkung. Dies ist in neuronalen Kernen üblich.
Dasjenige Outputneuron, welches sich am Ort der maximalen Erregung befindet, feuert als erstes und erregt die hemmenden Interneuronen. Diese hemmen alle ürigen Outputneuronen der Struktur, bevor diese selbst die Schwelle zur Auslösung eines Aktionspotentials erreichen. Somit feuert im Modul immer nur ein Outputneuron, die anderen schweigen.
Wohin führt der Output all der Projektionsneuronen in der tectalen x-y-Fläche?
Da es sich um einen motorischen Seitenwechselkern handelt, befindet er sich auf der motorischen Seite des alten Strickleiternervensystems bzw. des Neuralrohrs. Dort steigen die Signale auf den Axonen der Neuronen der Klasse 5 abwärts. Motorisch wirksame Signale erreichen über den Nucleus ruber die Motoneuronen der anzusteuernden Muskeln.
Doch der Output des tectalen, motorischen Seitenwechselkerns hat (mit einer Ausnahme!) keine motorischen Ziele. Ihm bleibt - im Nucleus ruber angekommen - nur der Weg zum Nucleus olivaris, der diese Signale über die Kletterfasern zum Cerebellum schickt. (Eine Ausnahme wird am Kapitelende vorgestellt.)
Somit gibt es im Cerebellum, genauer im Pontocerebellum, ein analog organisiertes Quadrat aus Purkinjezellen (genauer Purkinjegruppen), in denen der Output des tektalen motorische Seitenwechselkerns endet. Wir nennen dieses Gebiet das cerebellare Abbild des tectalen Seitenwechselkerns.
Der Zielort liegt mit Sicherheit im Pontocerebellum, weil das Spinocerebellum ausschließlich motorische Signale verarbeitet, die in unserem Fall jedoch nicht vorliegen.
Hier beginnt die Speicherarbeit des Pontocerebellums. Immer genau ein Projektionsneuron im tectalen motorischen Seitenwechselkern ist maximal erregt (denn es gibt nur ein globales Maximum). Das Maximum repräsentiert ein visuelles Objekt, welches mit beiden Augen fixiert wird. Vor allem aber repräsentiert dieses Maximum den Objektabstand und den mittleren Augenwinkel zum anvisierten Objekt.
Während die Stelle des schärfsten Sehens beim Fokussieren ein Abbild des Objekts auf der Netzhaut erzeugt, gelangen die visuellen Signale dieses Bildausschnitts über den visuellen Thalamus in den primären visuellen Cortex.
Von dort gibt es eine Projektion über die Brückenkerne ins Parallelfasersystem des Pontocerebellums. Wir unterstellen, dass die visuellen Signale der Retina in ihrer Gesamtheit über die Parallelfasern genau das cerebellare Abbild des tectalen Seitenwechselkerns erreichen. Wir unterstellen der Einfachheit halber, dass nur die Stelle des schärfsten Sehens eine Verbindung zum Cerebellum besitzt. Richtet man den Blick auf irgendein visuelles Objekt in der waagerechten Ebene, so müssen die Augen es mit dem Blick einfangen, wozu sie nach links oder nach rechts oder auch geradeaus schauen müssen. Je nach Abstand des Objekt sind die Winkel des linken und des rechten Auges meist unterschiedlich.
Nun beginnt die Analyse des Raumes in der Fläche (den Einfluss des Hebens und des Senken des Blicks haben wir hier noch nicht einbezogen, wir schauen also nur in der waagerechten Ebene).
Die Analyse erfolgt durch das Zusammenwirken zweier Algorithmen.
-
Im tectalen motorischen Seitenwechselkern wird ein Projektionsneuron
maximal erregt, dessen x-Koordinate den Fokussierwinkel ![]() und
dessen y-Wert den Blickrichtungswinkel
und
dessen y-Wert den Blickrichtungswinkel ![]() beim
Blick ins Unendliche angibt. Diese starke Erregung erreicht über den Nucleus
ruber und den Nucleus olivaris genau eine Purkinjegruppe im Gebiet des cerebellaren Abbilds des tectalen
Seitenwechselkerns. Diese Purkinjegruppe wird stark erregt und leitet den
Prägungsprozess ein, bei dem durch LTP und LTD die gerade aktiven
Parallelfasern eine starke Veränderung ihrer synaptischen Kopplungsstärke
erleiden und dauerhaft beibehalten.
beim
Blick ins Unendliche angibt. Diese starke Erregung erreicht über den Nucleus
ruber und den Nucleus olivaris genau eine Purkinjegruppe im Gebiet des cerebellaren Abbilds des tectalen
Seitenwechselkerns. Diese Purkinjegruppe wird stark erregt und leitet den
Prägungsprozess ein, bei dem durch LTP und LTD die gerade aktiven
Parallelfasern eine starke Veränderung ihrer synaptischen Kopplungsstärke
erleiden und dauerhaft beibehalten.
- Vom Gebiet des schärfsten Sehens erreichen alle visuellen Signale über den visuellen Thalamus den Cortex und von dort über die Brückenkerne genau die Parallelfasern aller Purkinjegruppen, die im Gebiet des cerebellare Abbilds des tectalen Seitenwechselkerns liegen. Alle dortigen Purkinjegruppen erhalten über die Parallelfasern das aktuelle Signalabbild des Gebietes des schärfsten Sehens.
- Aber nur genau eine dieser Purkinjezellen wird über die Kletterfasern aus dem tectalen motorischen Seitenwechselkern erregt. Nur diese Purkinjegruppe speichert das aktuelle Signalabbild aus dem Gebietes des schärfsten Sehens.
- Die übrigen Purkinjegruppen erhalten vom tectalen motorischen Seitenwechselkerns keinerlei Kletterfasererregung und können daher das aktuelle Signalabbild nicht speichern. Denn im tectalen motorischen Seitenwechselkerns unterliegt der Output einer neuronalen Konkurrenz. Hemmende Interneuronen sorgen dafür, dass sich nur der Output an der Stelle der Maximalerregung durchsetzt. Er vernichtet die vorhandene, deutlich geringere Erregung der übrigen Outputneuronen. Aber war der Blick in der zurückliegenden Zeit auf ein Objekt gerichtet, dessen Winkelwerte zu der schweigenden Purkinjegruppe gehört, so ist das Abbild des seinerzeit betrachteten Objekts in dieser Purkinjegruppe gespeichert.
- Da jedoch beim Sehen die Augen (unbewusst?) die gesamte Umgebung "abtasten", erstellt dieses System eine komplette Abbildung der Umwelt, die jedoch aus lauter kleinen Teilbildern besteht, die unser Gehirn jedoch als Gesamtbild wahrnimmt. Wandert der Blick hin und her, so bleibt dieses Gesamtbild unbewegt, lediglich der Aufmerksamkeitsfokus wandert mit einer Veränderung der Blickrichtung auf die Stelle der Gesamtabbildung, die durch die zugehörige Teilabbildung im Cerebellum repräsentiert wird. So erhalten wir den Eindruck, die Welt stehe still und nur unser Blick wandert hin und her (solange der Kopf nicht bewegt wird).
Damit gibt es zu jeder tatsächlich analysierten Blickrichtung ein visuelles Abbild im cerebellaren Abbild des tectalen Seitenwechselkerns. Dies ist der Grund dafür, warum wir die Umwelt so detailreich wahrnehmen und uns erinnern können, was wir wo in welcher Richtung gesehen haben, auch wenn wir kurzzeitig die Augen schließen. In unserem Unterbewusstsein, dem Pontocerebellum, genauer im cerebellaren Abbild des tectalen Seitenwechselkerns hat jede einzelne Purkinjegruppe einen Teilausschnitt der visuellen Umwelt abgebildet, sobald wir einmal in die zugehörige Richtung und Entfernung geschaut haben. Und dies tun wir automatisch. Wir suchen die Umgebung nach Objekten ab und speichern ihr visuelles Abbild in Sekundenschnelle in den Purkinjezellen des Pontocerebellums ab.
Und das inverse Pontocerebellum kann die abgespeicherten Objektabbildungen wieder im Bewusstsein aktivieren, indem es diese zurück zum visuellen Thalamus sendet, sobald die zugehörige Purkinjegruppe aktiviert wird. Dieser Prozess ist etwas komplizierter und wird hier nicht näher beleuchtet.
Zusammenfassend kann gesagt werden, dass der tectale motorische Seitenwechselkern die Grundlage für unsere Raumvorstellung liefert.
Ergänzung vom 20.10.2023:
Nun erinnern wir uns daran, dass es im Nucleus olivaris auch eine Signaldivergenz gibt. Das heißt, dass das aktuell aktive Neuron im tectalen Seitenwechselkern in mehrere Neuronen des Nucleus olivaris projiziert. Nehmen wir zunächst vereinfacht an, diese wären in einer Reihe hintereinandergeschaltet, so dass hier eine sequentielle Divergenz vorliegt. Dann könnten zu einer vorgegebenen Blickrichtung mehrere, verschiedene Bilder gespeichert werden. Es wäre also technisch möglich, verschiedene Objektabbilder zu speichern und später auch wiederzuerkennen. Dies könnten - beim Hamster, der den Heimweg sucht - markante Objekte am Wegesrand sein, die den Weg in den Hamsterbau markieren. Dies lässt den Schluss zu, dass diese neuronale Organisationsform bereits bei einfacheren Wirbeltieren vorhanden sein muss.
Bisher wurde jedoch eine Einschränkung gemacht: Die Augen durften nur in der waagerechten Ebene Ausschau halten. Diese Einschränkung lassen wir jetzt fallen. Nun untersuchen wir die Arbeitsweise des tectalen motorischen Seitenwechselkerns unter der Voraussetzung, dass der Blick der Augen auch nach oben oder nach unten gerichtet werden kann. Dann wird im tectalen motorischen Seitenwechselkern ein Raummodell der Umwelt geschaffen, bei dem jedem erkannten visuellen Objekt seine Raumkoordinaten zugeordnet werden können. Damit schafft dieses Modul die Grundlage unserer räumlichen Wahrnehmung.
Wir schauen uns noch einmal die Abbildung an, mit der wir dieses Kapitel begonnen haben.
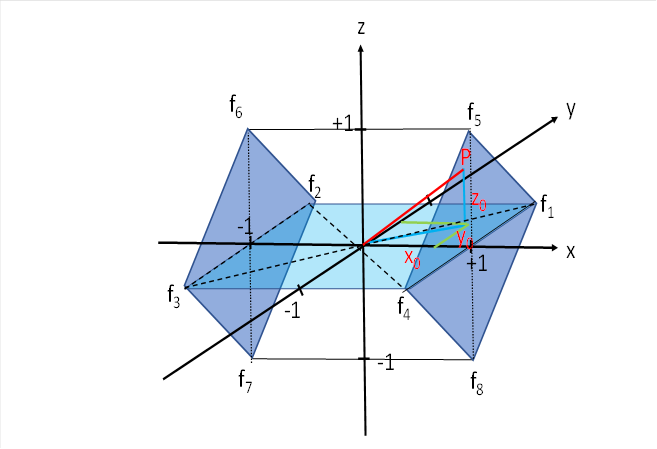
Bisher hatten wir nur das Quadrat betrachtet, welches sich in der x-y-Ebene befand. Für die Summe der vier Erregungen war im Kapitel "Das motorische Divergenzmodul mit seitlicher Signalausbreitung" bereits folgendes hergeleitet worden:
![]()
![]()
Wenn wir die Ebene verlassen und uns in den Raum begeben, müssen wir neben den Variablen x und y zusätzlich die Variable z berücksichtigen.
Da ein räumlicher Radiusvektor t aus dem Radiusvektor r der Ebene durch die zusätzliche Koordinate z ergänzt werden muss, lautet die neue Summenformel für die Erregungen in der Ebene folgendermaßen:
![]()
Hierbei gilt:
![]()
![]()
![]()
Setzt man dies in die obige Summenformel ein, so kann man den gemeinsamen Faktor vor die Summe ziehen:
![]()
Wegen
![]()
gilt auch:
![]()
Wegen
![]()
erhalten wir für die räumliche Erregungsfunktion im tectalen motorischen Seitenwechselkern die Gleichung
![]()
Nun ist besonders gut erkennbar, dass αp nur von der Variablen y und und αf nur von der Variablen x abhängt, wenn es um das globale Maximum geht.
Da dies der Anteil der Erregung ist, die von den Augenbewegungen in der Horizontalen verursacht wird, benennen wir ihn um und nennen ihn fortan fH. Dann gilt:
Die komplette Berechnung der partiellen Ableitungen von fH(x,y,z) ergibt folgende drei Gleichungen, die als Bedingungen für das Vorliegen eines globalen Maximums im Punkt P(x,y,z) erfüllt sein müssen:
- ![]()
- ![]()
- ![]()
Wir sehen, dass die ersten zwei Bedingungen identisch sind mit denen, die wir für das ebene tectale motorische Seitenwechselkern erhalten hatten.
Es fehlen nun noch vier Erregungsanteile, die von den Augenmuskeln geliefert werden, die für das Heben und Senken des Blick zuständig sind. Ihre Signale treffen in den oberen und unteren Ecken der zwei dunkleren, blauen Quadrate ein.
Wir betrachten aus der obigen Abbildung die zwei Quadrate, welches parallel zur y-z-Ebene liegen (dunkelblau). In seinen vier Ecken kommen die Augensignale an, die für das Heben und Senken des Blicks zuständig sind. Oben befinden sich die für das Heben des Blicks, unten für das Senken. Hierbei sind links die Signale des linken Auges zu finden und rechts die des rechten.
Wir betrachten nochmals die Abbildung zum tectalen, motorischen Seitenwechselkern.
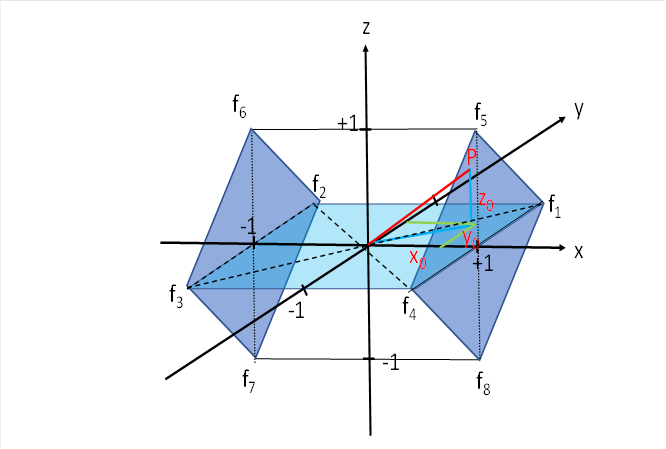
Wir müssen nun noch den Erregungsanteil fV für die Feuerraten f5 bis f8 berechnen, der die vertikalen Augenbewegungen ermöglicht. Wir bezeichnen mit t5 bis t8 die Radiusvektoren, welche die Abstande des Punktes P zu den vier betreffenden Ecken des Würfels darstellen, in denen diese vier Feuerraten eintreffen.
Prinzipiell gilt
![]()
Für die betreffenden Feuerraten gilt (siehe Kapitelanfang):
|
Musculus rectus |
Auge links |
Auge rechts
|
|
superior |
|
|
|
inferior |
|
|
Da die Feuerraten für das linke und das rechte Auge identisch sind, können wir bereits hier zusammenfassen.
![]()
Wir berechnen zunächst die Länge der Radiusvektoren, wobei wir gleich die Quadrate von ihnen benötigen.
Hierzu müssen wir den vier Ecken Punkte zuordnen und deren Koordinaten ermitteln, um die Länge der Radiusvektoren zuberechnen.
Im Punkt P5 treffe die Feuerrate f5 ein, in P6 die Feuerrate f6, zum Punkt P7 gehöre f7 und zu P8 gehöre f8.
Dann haben diese Punkte folgende Koordinaten:
|
Punkt |
x-Koordinate |
y-Koordinate |
z-Koordinate |
|
P5 |
+1 |
0 |
+1 |
|
P6 |
-1 |
0 |
+1 |
|
P7 |
-1 |
0 |
-1 |
|
P8 |
+1 |
0 |
-1 |
Der Abstand t eines Punktes mit den Koodinaten P(x,y,z) zu einem beliebigen Punkt mit den Koordinaten P(a,b,c) ergibt sich aus
![]()
Für die vier Punkte ergeben sich daher folgende Quadrate der Radiusvektoren:
![]()
![]()
![]()
Wir berechnen zunächst
![]()
![]()
![]()
![]()
Ebenso berechnen wir
![]()
![]()
![]()
![]()
Einsetzen in die Gleichung für fV liefert
![]()
![]()
![]()
Wir berechnen zunächst die Bedingungen für das Vorliegen eines Maximums für diese vertikal wirksamen Erregungsanteil, um uns darüber klar zuwerden, welchen Einfluss er hat.
Dazu berechnen wir die partiellen Ableitungen nach x, y und z und setzen sie gleich null.
![]()
Diese Gleichung ist nur erfüllt für
![]()
Daraus folgt sofort
![]()
Damit gilt für die Variable x die Extremwertbedingung:
![]() (Ableitung von fV nach x)
(Ableitung von fV nach x)
Wir berechnen die partielle Ableitung nach y und setzen sie gleich null, um eine weitere Bedingung für das Vorliegen eines Extremwertes zu erhalten.
![]()
Daraus folgt zwangsläufig folgende Extremwertbedingung:
![]() (Ableitung von fV nach y)
(Ableitung von fV nach y)
Als letztes berechnen wir die partielle Ableitung nach z und setzen sie gleich null, um die dritte Bedingugn für den Extremwert zu erhalten.
![]()
Diese Gleichung kann nur gleich null werden, wenn folgendes gilt:
![]()
![]()
![]()
![]()
![]()
Damit muss für das Vorliegen eines Extremwertes auch die dritte Bedingung erfüllt sein:
![]() (Ableitung von fV nach z)
(Ableitung von fV nach z)
Wir fassen zusammen:
Die Funktion fV(x,y,z) hat im Punkt P(x,y,z) genau dann und nur dann ein globales Maximum, wenn die folgenden drei Bedingungen (gleichzeitig) erfüllt sind
2. y=0
3. ![]()
Der Extremwert liegt also genau auf der z-Achse in der Höhe, die durch die dritte Gleichung vorgegeben ist. Mit wachsendem z wird der Winkel β größer, mit abnehmendem z wird dieser Winkel kleiner.
Oder umgekehrt: Je größer z ist, um so größer muss der Winkel β werden, damit der Extremwert wieder im Punkt z liegt.
Wir erinnern uns jedoch, dass diese Gleichungen nur den Erregungsanteil der Augenmuskel gelten, die für die vertikalen Augenbewegungen (nach oben und nach unten) enthalten.
Normalerweise werden die Sehnenorgane der Augenmuskel sowohl eine Beitrag zur Horizontalerregung fH als auch einen Beitrag zur Vertikalerregung fV leisten, da in den meisten Fällen der Blick ja nicht starr geradeaus, sodern sowohl etwas seitlich als auch nach oben oder unten ausgerichtet ist.
In diesen Fällen überlagern sich beide Erregungskomponenten und addieren sich. Dadurch verschiebt sich das Maximum. Wegen der Konkavität der Teilfunktionen und des Überlagerungsergebnisses liegt das neue Maximum auf einer Verbindungslinie der ursprünglichen zwei Maxima, die jede der Teilfunktionen hatte. Leider ist diese Verbindungslinie keine Gerade, sondern gekrümmt. Daher ist die Position des neuen Extremwertes nur sehr schwer zu berechnen.
Wir erinnern uns: Der horizontale Anteil der Erregung betrug
![]() .
.
Die zugehörigen Bedingungen für ein Maximum im Punkt P(x,y,z) waren
- ![]()
- ![]()
- ![]() .
.
Der vertikale Anteil der Erregung betrug
![]() .
.
Die zugehörigen Bedingungen für ein Maximum im Punkt P(x,y,z) waren
- x=0
- y=0
- ![]()
Jede der zwei Teilerregungen besitzt bereits ein Gradientenfeld, welches nicht kugelsymmetrisch ist, sondern durch die Parameter der Augenwinkel etwas verzerrt ist.
Während der horizontale Anteil ein Erregungsfeld darstellt, welches im Punkt PH(x,y,0) ein globales Maximum besitzt, befindet sich das Erregungsmaximum des vertikalen Anteils genau auf der z-Achse in der Höhe z, also im Punkt PV(0,0,z).
Durch die Überlagerung beider Anteile wird das Maximum in Punkt PH(x,y,0) in Richtung des Punktes PV(0,0,z) gezogen, jedoch nicht geradlinig, sondern entlang der aktuellen Gradienten. Diese sind jedoch (meist) gekrümmt. Entlang der geradlinigen Verbindung beider Punkte existiert wegen der strengen Konkavität beider Anteile auch ein Maximum, jedoch ist es nur ein lokales entlang dieser Geraden. Das absolute Maximum liegt irgendwo in der Nähe dieser Linie, jedoch ist seine korrekte Bestimmung extrem schwierig. Eine Ermittlung der drei Augenwinkel αF, αP und β durch Nullsetzen der partiellen Ableitungen erscheint dem Autor nach vielen Versuchen als unmöglich. Mögen andere den Versuch wagen.
Als Ergebnis bleibt jedoch festzuhalten:
Im räumlichen tectalen motorischen Seitenwechselkern repräsentiert die Maximalerregung eines der vielen Outputneuronen genau einen Raumpunkt, der von den Augen anvisiert wird. Die Maximalerregung entsteht im Modul einerseits durch den Input der beteiligten acht geraden Augenmuskeln der zwei Augen, andererseits durch die räumliche Ausbreitung und Überlagerung dieser Erregungen im dreidimensionalen Raum des Moduls. Die Signaldämpfung während der Ausbreitung nimmt quadratisch mit der Entfernung zu. Dadurch ergibt sich eine streng konkave Überlagerungsfunktion mit einem eindeutigen globalen Maximum im Modul.
Die Abbildung des Umgebungsraumes im tectalen motorischen Kern wird übertragen ins Cerebellum. Hierbei muss jedoch die räumliche Abbildung in eine Abbildung in der Fläche übertragen werden, denn die Purkinjezellen bilden eine Fläche. Der Würfel (siehe Abbildung) muss also in Scheiben geschnitten werden, die anschließend nacheinander angeordnet werden. Jede Scheibe repräsentiert eine Raumhöhe, wir wollen sie als Höhenscheibe bezeichnen. Da die Fläche nach oben und unten schmaler wird (der Würfel steht gedreht im Raum), werden die Höhenscheiben auch schmaler. So hat das repräsentierte Gebiet im Cerebellum die Form eines länglichen Rhombus, der viel schmaler ist als seine Gesamtlänge.
Diese Zerlegung in Scheiben ist auch der Grund dafür, warum die neuronale Repräsentation eines bewegten Objekts im Raum im Cerebellum, aber auch im nachfolgenden Hippocampus - plötzlich Sprünge aufweist. Bewegt sich das Objekt so, dass ein Übergang von einer Scheibe in eine andere erfolgt, so springt das Maximum plötzlich von der einen in die andere Scheibe. Hierbei tritt eine Diskuntinuität auf.
Jedes dieser Gebiete wird über eine aufsteigende Projektion kopfwärts gesendet und erreicht die erste Etage des frühen Strickleitersystems, wo es letztlich im Hippocampus landet. Dort finde man ebenfalls die hintereinander angeordneten Höhenscheiben, von denen die für die waagerechte Ebene die größte Breite hat.
Bewegt sich nun ein visuelles Objekt in der Ebene hin und her, beispielsweise eine schwimmende Ente auf einer Wasserfläche, so wird in der zugeordneten Höhenscheibe die Abbildung dieser Ente ebenfalls hin- und herbewegt. Entlang des Weges der Ente wird immer genau ein Hippocampusneuron maximal erregt sein, weil das zugehörige Neuron im tectalen motorischen Seitenwechselkern maximal erregt ist und die Erregung über das Cerebellum den Hippocampus erreicht. Dieser enthält eine Abbildung des visuellen Umgebungsraumes, und dasjenige Neuron, welches durch die Signale der Sehnenorgane der Muskeln im tectalen Divergenzmodul maximal erregt ist, repräsentiert das angepeilte visuelle Objekt - in unserem Fall die Ente. Verlässt das Abbild des Objektd durch eine Höhenänderung die aktuelle Höhenscheibe, so springt das Erregungsmaximum in die nächste aktuelle Höhenscheibe - zeigt also ein unstetiges Verhalten.
Man sollte diese Abbildung des tectalen motorischen Seitenwechselkerns nicht als Navigationsgerät auffassen, denn es ist immer eine sehr schnell vergängliche Abbildung des momentanen Zustandes der Augenmuskeln. Sie ist daher eher eine Karte der Augenrichtungen.
Dennoch wird hier bereits das Phänomen auftreten, dass die Erregung sich beim Bewegen des Objekts - beispielsweise der Ente - immer entlang von Sechsecken ausbreitet. Genauer gesagt sind es jedoch Dreiecke, die ihrerseits Sechsecke bilden.
Warum ist das so?
Die vielen Tausend Axone, die die Signale vom Cerebellum zum Hippocampus befördern, können als gerade Rohrleitungen konstanten Durchmessers interpretiert werden. Und diese ordnen sich bei Platzmangel so an, dass die Rohre versetzt übereinander liegen und dabei im Querschnitt eine Art Wabenform aus lauter kleinen Dreiecken entsteht. Somit hat die beobachtete Wabenform im Hippocampus im Gebiet der Repräsentation der Augenrichtungen ihre Ursache in der platzsparenden Anordnung der Axone, die später beim Auffächern in der Fläche diese wabenähnliche Form beibehalten. Daher muss man nicht nach neuronalen Netzen suchen, die durch -Lernvorgänge- zur Wabenform der aktiven Signale führen. Es gibt im Nervensystem oft einfache Erklärungen.
Auch die Entdeckung von Spiegelneuronen ist so ein Fall. Wenn eine Ente eine andere betrachtet, die auf dem Wasser herumschwimmt, dann ist der Weg der sich bewegenden Ente im Hippocampus der beobachtenden Ente durch maximalerregte Neuronen zu erkennen, die den Weg der sich bewegenden Ente nachzeichnen. Hier liegen keine Spiegelneuronen vor. Wenn Neuronen feuern, gibt es immer einen erklärbaren Algorithmus, warum sie feuern. Sie sind nicht in der Lage, Handlungen des Gegenüber -nachzuvollziehen-. Überhaupt sollten wir uns abgewöhnen, Neuronen eine Intelligenz zuzusprechen und Formulierungen zu verwenden wie: Das Neuron erkennt --
Neuronale Verschaltungen und Rezeptorinput zwingen bestimmte Neuronen, unter bestimmten Bedingungen maximal zu feuern. Intelligent ist dann nicht das feuernde Neuron, sondern das Lebewesen, in dessen Nervensystem dies alles stattfindet. Intelligenz ist eine Systemeigenschaft, sie lässt sich nicht einzelnen Neuronen zuweisen.
Am Ende dieses Kapitels beantworten wir die Frage, welche Signale die Augenlinse in ihrer Form so verändern, dass sie das anvisierte Objekt auf der Netzhaut scharf abbildet.
Der Muskel, dessen Kontraktion die Krümmung der Augenlinse und damit ihre Brechzahl vergrößert, ist der Ziliarmuskel, der die Linse ringförmig umgibt und an flexiblen Zonulafasern im Auge befestigt ist. Er besteht aus drei Muskeln: dem Brückeschen Muskel, dem Iwanoffschen Muskel und dem Müllerschen Muskel. Bei Kontraktion des Brückerschen Muskels wird die Brechkraft der Linse vergrößert, dadurch wird die Nahakkumodation ermöglicht. Der Müllersche Muskel bewirkt ebenfalls eine Vergrößerung der Brechkraft. Der motorische Gegenspieler dieser zwei Muskeln ist der Iwanoffsche Muskel. Wenn er kontrahiert, wird die Brechkraft der Linse kleiner, so wird eine Sehschärfe beim Sehen in die Ferne eingestellt.
Wir benötigen für die Steuerung der Akkomodation zwei Muskelsignale: eines versorgt den Brückerschen und den Müllerschen Muskel mit Kontraktionssignalen, das andere Signal ist invers dazu und ist für den motorischen Gegenspieler, den Iwanoffschen Muskel bestimmt.
Dieses Signalpaar entsteht ganz nebenbei im tectalen motorischen Seitenwechselkern.
Wir schauen uns nochmals die zugehörige Abbildung an.
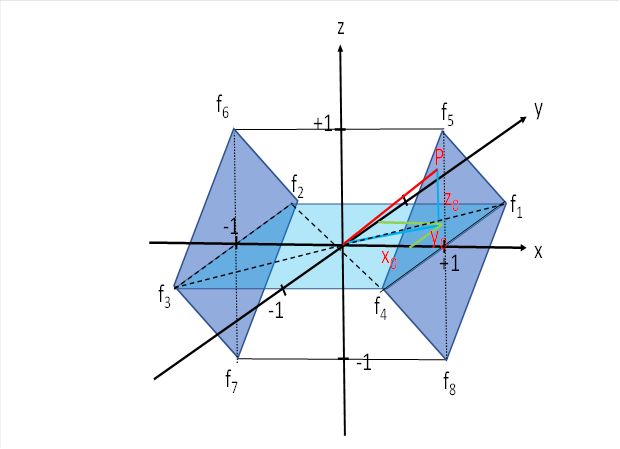
Wir benötigen für jedes Auge zwei Signale. Folgende Signale sind dafür erforderlich:
![]() rechtes Auge, muscilus rectus
lateralis, Blick nach rechts
rechtes Auge, muscilus rectus
lateralis, Blick nach rechts
![]() linkes Auge, musculus rectus medialis,
Blick nach rechts
linkes Auge, musculus rectus medialis,
Blick nach rechts
![]() rechtes Auge, musculus rectus medialis,
Blick nach links
rechtes Auge, musculus rectus medialis,
Blick nach links
![]() linkes Auge, musculus rectus lateralis,
Blick nach links
linkes Auge, musculus rectus lateralis,
Blick nach links
Wir erinnern uns an folgende Zusammenhänge:
![]()
![]()
Dann gelten auch folgende Gleichungen:
![]()
![]()
![]()
![]()
Wir betrachten nur die Signale in der waagerechten Ebene (hellblau). Zuerst bilden wir den Quotienten aus f1 und f2.
![]()
Dieser Feuerratenquotient enthält nur noch den Fokussierwinkel αF. Dieser Winkel ist der Unterschied zwischen der Augenrichtung des linken und des rechten Auges, er ist zum Fokussieren nötig. Daher eignen sich die zwei Größen f1 und f2 zur Ansteuerung der Muskeln für die Brechkrafteinstellung des Auges. Für den Brückerschen und den Müllerschen Muskel ist die Feuerrate f1 geeignet, und zur Ansteuerung des Gegenspielers, des Iwanoffschen Muskels, ist die Feuerrate f2 zuständig.
Denn f1 nimmt mit wachsendem Winkel αF zu, während f2 mit kleiner werdendem Winkel αF zunimmt. Und da es bei der Muskelspannung nur auf den Quotienten aus beiden ankommt (Extremwerte bleiben unverändert bei Multiplikation der Ausgangsfunktionen mit Konstanten), fällt der Winkel αP aus der Betrachtung heraus.
Damit ist das Feuerratenpaar (f1;f2) für die Ansteuerung des Ziliarmuskels mit seinen drei Komponenten geeignet. Und dieses Signal kann sowohl für das linke als auch für das rechte Auge verwendet werden und steuert so die entfernungsabhängige Akkomodation der Augenlinse.
So erfüllt das tectale motorische Seitenwechselkern auch die Aufgabe der Scharfeinstellung der Augenlinse.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan