Gehirntheorie des Menschen
ISBN 978-3-00-068559-0
Monografie von Dr. rer. nat. Andreas Heinrich Malczan
5. Die Basalganglien – ein Verzögerungsmodul
Neben den abwärts projizierenden Neuronen der Klasse 5 gab es im alten Strickleitersystem bereits die abwärts projizierenden Neuronen der Klasse 6. Während die Axone der Klasse-5-Neuronen auf der motorischen Seite abwärts zogen, konnten die Axone der Mittelwertneuronen der Klasse 6 sowohl auf der sensorischen als auch auf der motorischen Seite abwärts ziehen, denn sie waren sowohl in motorischen als auch in sensorischen Kernen vorhanden.
Ebenso gab es die Neuronen der Klasse 4, die den aufsteigenden Input von unten empfangen konnten, aber die Neuronen der Klasse 1 konnten dies auch. Während es die Klasse-4-Neuronen nur auf der sensorischen Seite gab, existierten die Neuronen der Klasse 1 auf beiden Seiten, der sensorischen und der motorischen.
Die Neuronen der Klasse 6 und der Klasse 1 gehören zum Mittelwertsystem und sind auch heute in allen Wirbeltieren zu finden. Aus dem Mittelwertsystem entwickelte sich das Basalgangliensystem.
Die Basalganglien sind ein wichtiges Subsystem des Gehirns, über dessen Funktion und Wirkungsweise viele unterschiedliche Ansichten verbreitet sind. Die Erklärung ihrer signalverarbeitenden Funktion in der gegenwärtigen Fachliteratur bedarf einer dringenden Korrektur!
Die zentrale Struktur der Basalganglien ist die dopaminerge Substantia nigra pars compacta und die dazugehörige Area tegmentalis centralis (VTA). Sie sind gemeinsam der dopaminerge Mittelwertkern des Gehirns. Ursprünglich gab es wohl auch diesen Mittelwertkern in jedem Segment, doch durch die neuronale Konkurrenz über die laterale Hemmung blieb nur der Kern im siebenten Segment übrig, dort kamen ja auch alle Signale aus dem Rumpf an.
Zur Entstehung trug der Umstand bei, dass alle sensorischen und motorischen Kerne eine Rückprojektion von den Mittelwertkernen erhielten. Zunächst war die Projektion immer auf ein Segment beschränkt, doch auch die Mittelwertkerne überwanden mit ihren Signalen die Segmentgrenzen. Damit begann der neuronale Wettbewerb, der erst beendet wurde, als ein Mittelwertsystem (pro Transmitter) die Oberherrschaft gewonnen hatte und die entsprechenden Mittelwertzentren sich in den übrigen Segmenten zurückbildeten. In den ersten sechs Segmenten setzten sich die abhängigen Rezeptoren durch und verdrängten die übrigen Rezeptorenarten, aber auch die meisten Mittelwertkerne. Eine Ausnahme bildet das erste Segment, welches als ältestes System – von ihm stammen alle segmentierten Lebewesen ab – sehr konservativ war und weiteren Veränderungen häufig widerstehen konnte. Durch die Auffaltung der Modalitäten befindet es sich in der Temporalschleife, dem späteren Temporallappen.
So blieb im siebenten Segment (unter anderem) das dopaminerge Mittelwertzentrum bestehen. Die Area tegmentalis centralis ist derjenige Anteil dieses Mittelwertkerns, der Input aus dem olfaktorischen, später limbischen System empfängt. Die Substantia nigra pars compacta dagegen erhält den übrigen Input. Wahrscheinlich führte der neuronale Wettbewerb zur Atrophie der dopaminergen Mittelwertzentren in den Rumpfsegmenten. Dies wäre noch zu prüfen.
Die Spezialisierung des siebenten Segments auf ein dopaminerges Mittelwertzentrum hatte zur Folge, dass jedes Kopfsegment die Signale seiner sensorischen und motorischen Kerne über Neuronen der Klasse 6 in dieses dopaminerge Mittelwertzentrum sandte. Wir sollten jedoch bedenken, dass diese Neuronen ihren Input in sensorischen Kernen und im sensorischen Cortex aus der Schicht 3 empfingen. In motorischen Kernen und im motorischen Cortex waren die Neuronen der Klasse 5 die Inputlieferanten für die Neuronen der Klasse 6. Letztere projizierten zum dopaminergen Zentrum des siebenten Segments. Offenbar wurden sie durch einen Marker gesteuert, der von diesem Zentrum in die Umgebung diffundierte. Die Neuronenklasse 5 dagegen folgt offenbar einem motorischen Marker, der zudem segmentspezifisch ist. Unterscheidbar sind die Axone dieser zwei Klassen nur sehr schwer, da sie beide abwärts ziehen.
Die olfaktorischen bzw. limbischen Signale des ersten Segments erreichen hierbei die VTA, die übrigen Segmente projizieren alle in die Substantia nigra pars compacta, die direkt benachbart liegt. Sie ist dennoch nach Modalitäten wohlgeordnet, da jedes Kopfsegment andere Modalitäten verarbeitete. Wir unterstellen daher, dass auch der dopaminerge Kern nach Modalitäten geordnet war. Erkennbar wird dies auch daran, dass unterschiedliche Lobi des Gehirns in unterschiedliche Abschnitte dieses Kerns projizieren.
Speziell auch das erste Segment, der spätere Cortex, projizierte mit allen nichtolfaktorischen Signalen in das siebente Segment zur Substantia nigra pars compacta.
Nun projiziert jeder Mittelwertkern zurück zum Ort seiner Signalherkunft. Daher gibt es eine Rückprojektion der Substantia nigra pars compacta und der VTA in Richtung der Signalherkunft. Diese endet jedoch – wie cytoarchitektonische Untersuchungen belegen - meist an den hemmenden Interneuronen der Zielstrukturen, die die ursprünglichen Signale geliefert haben.
Welche Veränderungen im Reaktionsverhalten bewirkte die Rückprojektion?
Es war der Zeitunterschied. Die rückkehrenden Signale waren zeitverzögert. Hatte sich in der Zwischenzeit nichts geändert, so wurde der Output mit seiner eigenen Feuerrate gehemmt. Nur wenn sich etwas veränderte – also bewegte – blieb ein Restsignal übrig, jedoch nur, wenn das (erregende) Gegenwartssignal stärker war als das hemmende Vergangenheitssignal. So konnten Bewegungen visuell erkannt, gerochen, ertastet werden. Auch Eigenbewegungen wurden wahrnehmbar. Die hemmende Rückprojektion begründete im Striatum der Basalganglien die sogenannte Matrix. Deren Axone werden von Dopamin gehemmt.
Es gab jedoch – und dies viel früher – auch eine erregende Rückprojektion aus dem dopaminergen Mittelwertkern des siebenten Segments.
Die erregende dopaminerge Projektion erfolgte zu den Striosomen, aus denen die Basalganglien zunächst bestanden. Sie projizierten hemmend in den Nucleus ruber und erzeugten dort eine zeitsensitive Differenzabbildung zur Bewegungsanalyse. Ihre Entstehung und ihre Funktion wurden in meiner vorherigen Monografie beschrieben und werden hier übersprungen.
Die hemmende Rückprojektion bildete sich in späterer Evolutionszeit. Anfänglich hemmte sie zeitverzögert die Signalausgabe im Cortex, wenn sich in der Zwischenzeit nichts geändert hatte.
Da es jedoch günstiger war, wenn die bisherige erregende Projektion erhalten blieb und die bisherigen Reaktionen im Rückenmark auslöste, wurde das Differenzsignal irgendwann nicht mehr im Cortex gebildet, sondern führte zur Bildung von eigenständigen Projektionsneuronen. Daher sind die Basalganglien ein Kern mit eigenem Output.
Wie entstand dies?
Die hemmende dopaminerge Rückprojektion wurde mit der Zeit spezifisch. Jedes Cortexneuron, jedes Outputneuron eines sensorischen oder motorischen Kerns in den Segmenten 1 bis 6 projizierte in genau ein Neuron der Substantia nigra pars compacta. Dieses Neuron schaltete den Transmitter von Glutamat auf Dopamin um und sandte sein Axon in die Herkunftsstruktur des Signals. Dort dockte sein Axon an einem hemmenden Interneuron an.
Diese hemmenden Interneuronen, die den dopaminergen Input von der Substantia nigra pars compacta bzw. von der VTA empfingen, sonderten sich räumlich von der ursprünglichen cortikalen Struktur ab und bildeten einen eigenen Kern. Im ersten, cortikalen Segment entstand so das Striatum unterhalb der Cortexrinde. Seine hemmenden Projektionsneuronen sind Abkömmlinge der hemmenden Interneuronen des Cortex. Der hemmende Output dieses neuen Kerns konnte nun mit den ursprünglichen Cortexsignalen und den Signalen der übrigen Segmente überlagert werden. Diese Überlagerung fand teilweise im Nucleus ruber statt, durch den ja der gesamte motorische Output des Urhirns zum Zweck der motorischen Steuerung des Körpers zu den Motoneuronen des Rumpfes zog. Ein Teil der Striatumneuronen – genauer die Striosomenneuronen – projizierten hemmend in den Nucleus ruber. Diese Axone sind ein wichtiger Bestandteil des Tractus tegmentalis centralis.
Die GABA-egen Matrixneuronen des Striatums projizierten abwärts und erreichten (auf der motorischen Seite) den motorischen Kern der zweiten Etage, der selbst eine komplette Kopie des sensorischen Kerns dieser Etage von den Neuronen der Klasse 3 geliefert bekam, die erregend war. Dort überlagerten siche diese Signale in einer Punkt-zu-Punkt-Abbildung. Während der sensorische Kern dieser Etage zum sensorischen Thalamus wurde, entwickelte sich aus dem motorischen Kern der motorisch (ventrale) Thalamus.
Diejenigen Signale des sensorischen Thalamus, die zu den sensorischen Cortexgebieten projizierten, durchliefen danach die Basalganglien und kehrten nach dem Durchlaufen der Basalganglien hemmend zum motorischen Thalamus zurück. Dieser empfing vom sensorischen Thalamus den gleichen, jedoch erregenden Input über die Neuronen der Klasse 3. Die punktweise Überlagerung beider Signalarten erzeugte eine zeitsensitive Differenzabbildung, die auf Bewegungen reagierte. Diese wurde wieder dem Cortex zugestellt und erreichte dort neue Assoziationsgebiete zur Bewegungs- und Richtungserkennung. Der motorische Thalamus entspricht für visuelle Modalitäten dem Pulvinar. Er empfängt über Kollateralen eine komplette erregende Kopie des primären visuellen Thalamus, die mit den hemmenden Striatumsignalen überlagert wird und eine zeitsensitive Differenzabbildung erzeugt, die Bewegungen visueller Art erkennt. Der Output erreicht neue, sekundäre cortikale Assoziationsgebiete, die der Analyse von Bewegungen und Orten dienen.
Damit gab es Überlagerungsgebiete im motorischen Thalamus. Wir wollen ihn als sekundären Thalamus bezeichnen. Der primäre Thalamus empfängt – wie auch der primäre Cortex (mit Ausnahme des Frontalcortex) den Input von Rezeptoren.
Die dopaminergen Signale waren gegenüber den Ursprungssignalen, von denen sie durch Transmitterumschaltung gewonnen wurden, zeitverzögert. Der Umweg über die Substantia nigra pars compacta bzw. die VTA hatte sie um etliche Millisekunden verzögert. Nach der Umschaltung auf GABA erreichten sie die Zielstrukturen im sekundären Cortex zwecks Überlagerung mit den ursprünglichen Signalen mit einer gewissen Zeitverzögerung.
Damit wurde eine Bewegungserkennung möglich. Hatte sich in der Zwischenzeit nichts geändert, so löschte das hemmende Vergangenheitssignal aus dem Striatum das erregende, unverzögerte Gegenwartssignal aus. Wenn jedoch das erregende Gegenwartssignal an Stärke zugenommen hatte, konnte das alte Vergangenheitssignal, das ja etliche Millisekunden zuvor entstanden war, dieses nicht mehr komplett auslöschen. Es blieb ein Restsignal übrig, dessen Stärke die Signaländerung verschlüsselte. Über den Nucleus ruber erreichte es die Motoneuronen und löste Reaktionen aus, die zuvor unmöglich waren.
So dienten Basalganglien der Bewegungserkennung. Überall dort, wo ein Objekt, z. B. ein Beutetier, sich bewegte, konnte seine Bewegung mittels der Basalganglien gerochen, ertastet, vom Seitenliniensinn erfasst und gesehen werden. Jede Modalität war eingebunden, denn jedes Kopfsegment und jedes Rumpfsegment sandte seine Signale in Richtung Cortex und in die sieben Kopfsegmente, wo die Bewegungsanalyse sogar nach Modalitäten getrennt analysiert werden konnte.
Und weil es günstiger war, die ursprünglichen, ungehemmten Signale ebenfalls zu behalten, wurden diese Originalsignale aufgeteilt. Eine komplette Signalkopie erreichte einen neuen Thalamuskern, der auch als ventraler Thalamus bezeichnet wird. Dort liegt eine komplette Kopie der Cortexsignale vor. Genutzt werden dazu die Neuronen der Klasse 3. Sie projizieren in jedem Segment von der sensorischen zur motorischen Seite. Da die gleichen Signale zum Cortex aufsteigen, ebenfalls von der sensorischen zur motorischen Cortexseite wechseln und wieder absteigen, kommen sie wieder an diesem ventralen Thalamus vorbei. Dort münden auch die zeitverzögerten und hemmenden Signale aus der Matrix des Striatums ein und löschen all die Gegenwartssignale aus, die sich in der Zwischenzeit nicht geändert hatten. Nur Bewegungen oder Signalstärkeveränderungen blieben übrig.
So besaß dieser Kern die zeitsensitive Differenzabbildung und konnte sie wiederum erregend an den Cortex zur weiteren Auswertung senden.
Da die Funktionsweise der Basalganglien vorwiegend auf der Hemmung von Mittelwertsignalen beruht, also letztlich eine Signalinversion darstellt, wird hierbei auch eine Monotonieumkehr realisiert. Ein maximumcodiertes Signal wird durch eine Monotonieumkehr jedoch minimumcodiert. Daher ist eine zweite Signalinversion nötig. Diese wird vom Globus pallidus realisiert. Dadurch wird der invertierte Output wieder maximumcodiert. In den Basalganglien wird also jedes maximumcodierte Signal zweimal invertiert, eimal in der Matrix des Striatums und einmal im Globus pallidus. Dies ist ein sehr starkes Indiz für die Richtigkeit meiner Theorie der Extremwertcodierung und der Signalinversion.
Für den motorischen Bereich ist die Schaltung der Basalganglien in der folgenden Abbildung von mir dargestellt.
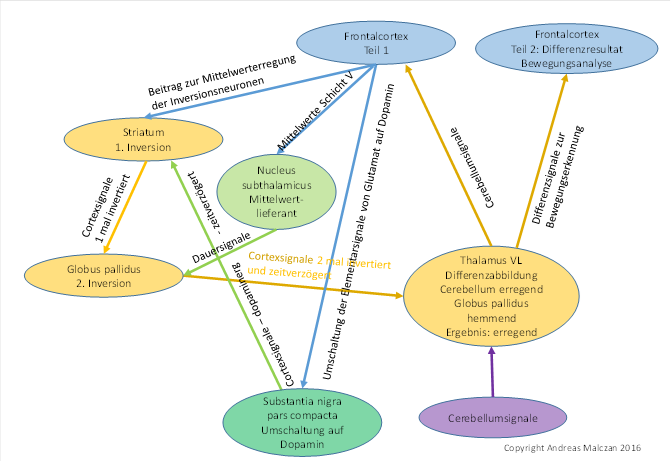
Abbildung 37: Basalganglienmodul und sein Verschaltungsprinzip für motorische Signale
Die schrittweise Entwicklung der Basalganglien habe ich in meiner Monografie „Gehirntheorie der Wirbeltiere“ ausführlich dargestellt. Es wird Zeit, dass sich unsere führenden Neurologen von den veralteten und unrichtigen Ansichten zu den Basalganglien trennen, die noch heute durch alle neurologischen Fachbücher geistern. Insbesondere empfehle ich das Studium der Aufteilung des Striatums in die Striosomen und die Matrix, die in meiner vorherigen Monografie ausführlich dargestellt wird. Hier muss die aktuelle Auffassung von der Funktionsweise der Basalganglien komplett neu überdacht werden.
Nun fehlen noch drei wichtige Entwicklungen im Wirbeltiergehirn. Dies sind
1. Die Myelinisierung der Projektionsaxone
2. Die Aufspaltung der Modalitäten
3. Die Signaldivergenz
Es wäre müßig, über die Bedeutung der Bildung des Myelins für die neuronale Signalweiterleitung zu schreiben.
Daher folgt nun die Beschreibung des Verzögerungsmoduls zum Bewegungssehen im Hell-Dunkel-Format. Die Schaltung des Verzögerungsmoduls für sämtliche Modalitäten ist identisch, deshalb behandeln wir hier beispielhaft die visuelle Erkennung von Bewegungen durch das Hell-Dunkel-Modul. Es gestattet die Bewegungsanalyse von Objekten, die mit dem Hell-Dunkel-Modul des visuellen Cortex analysiert werden. Die visuellen Farbmodule arbeiten nach dem gleichen Prinzip.
5.1 Das visuelle Verzögerungsmodul der Basalganglien
Im Kapitel 4.2.5. über das Helligkeitsmodul mit räumlicher Signalausbreitung hatten wir untersucht, wie Orientierungssäulen im visuellen Cortex es ermöglichten, den Anstiegswinkel einer hellen Geraden und ihren Nullpunktabstand über eine Maximumcodierung zu verschlüsseln. Damit wurde eine wichtige Vorleistung für die Formerkennung von dunklen Objekten erbracht, denn die erkannten Linienelemente erlaubten es, den Umriss eines dunklen Objektes zu erkennen. Nach dem gleichen Prinzip erfolgte die Erkennung von hellen Objekten. Anschließend wurde das Analyseprinzip auf die Helligkeitsmodule und die Farbmodule des menschlichen Gehirns übertragen.
Je vier benachbarte retinale und magnocellulare Ganglienzellen eines Auges versorgen (über den visuellen Thalamus) ein würfelförmiges Hell-Dunkel-Modul im Schichtsystem S4-Dunkel-On//S3-Dunkel-On//S4-Hell-On. Sie bilden ein Quadrat – jedoch nur theoretisch. Ebenso könnten sie einen Rhombus bilden, bei dem jede Seite eine andere Länge hat. Das mathematische Prinzip der Extremwertbildung durch Überlagerung konkaver Funktionen bleibt auch dann bestehen.
Ebenso versorgen je vier am gleichen Retinaort angeordnete parvocellulare Ganglienzellen vom Typ Blau die darüber angeordnete Inputschicht S4-Blau. Es folge eine Outputschicht S3-Blau, die jedoch auch Input aus der darüber angeordneten Inputschicht S4-Grün entgegennehmen kann. Über der Schicht S4-Grün folgt die Outputschicht S3-Grün, die auch Input von der darüber liegenden Inputschicht S4-Rot empfängt. Über S4-Rot liegt die Outputschicht S3-Rot, die auch Input von der darüber gelegenen Inputschicht S4-Blau annehmen kann. Die ganz oben befindliche Outputschicht S3-Blau konnte im Verlauf der Evolution degenerieren, sie wurde durch die Entwicklung der Signalüberlagerung überflüssig.
Damit gab es in diesem quaderförmigen Cortexmodul sechs recht dünne Inputschichten, zwei für Hell-On und Hell-Off, zwei für den Farbinput Rot und Grün sowie zwei Schichten für die Farbe Blau, Letztere je eine untere und eine obere Schicht.
Und es gab eine Outputschicht für die Signalart Helligkeit sowie drei für die Mischfarben Blau/Grün, Grün/Rot und Rot/Blau.
Jede dieser Orientierungssäulen, die ein Retinaquadrat analysierten hinsichtlich Helligkeit und Farbe, besaß in seinen insgesamt vier recht dicken Outputschichten viele Tausend oder Hunderttausend Outputneuronen.
Wir haben gelernt, dass die Outputschichten 3 – auch wenn sie in Subschichten „zerfallen“ waren, ihren Output über zwei Richtungen weitergaben. Nach oben gab es die Schicht 2 – die dann (wahrscheinlich) auch Subschichten bildete, die den Subschichten vom Typ 3 zugeordnet waren. Sie projizierten zu anderen Cortexgebieten.
Ebenso gab es den Signaltransport nach unten. Anfänglich übernahm die Schicht 6 die Signale der Schicht 3 und versorgte die Mittelwertstrukturen im siebenten Segment. So erhielt auch die dopaminerge Substantia nigra diese Signale.
Als jedoch in den Schichten des Typs 3 die Neuronenanzahl stark zunahm und diese Schicht sowie die aus ihr hervorgehenden Subschichten immer breiter und dicker wurden, machte es keinen Sinn, diese Signale von großen Mittelwertsignalen zusammenfassen zu lassen. Viel günstiger war es, wenn diese Signale unverfälscht zur Substantia nigra ziehen konnten. Dort wurden sie auf den Transmitter Dopamin umgeschaltet. Nun zogen sie zurück zur Matrix des Striatums. Dort wurde auf GABA umgeschaltet und die Signale wieder zum ventralen Thalamus geschickt.
Dort kam jedoch auch der Output des primären visuellen Cortex an. Denn dieser projizierte in sekundäre Felder auf der motorischen Seite des Occipitallappens. Dessen absteigende Signale durchquerten ebenfalls diesen ventralen Thalamus.
Damit standen im ventralen Thalamus für den gesamten primären visuellen Cortex zwei Signalarten zur Verfügung:
- Jedes Outputsignal einer jeden Orientierungssäule endete an einem eigenen Neuron. Es war erregend.
- Jedes dieser Signale erreichte zeitverzögert über die Wegstrecke Substantia nigra pars compacta-Matrix des Striatums ebenfalls den ventralen Thalamus. Es war hemmend.
Damit gab es für jedes einzelne Outputneuron im ventralen Thalamus das erregende Gegenwartssignal und das daraus gewonnene, hemmende Vergangenheitssignal. Beide endeten am genau einem Outputneuron des ventralen Thalamus. Der Output dieses Überlagerungsneurons trat nur dann auf, wenn das Vergangenheitssignal das Gegenwartssignal nicht komplett auslöschen konnte.
Da jedes Outputneuron genau zu einer Orientierungssäule gehörte, konnte das Vergangenheitssignal das Gegenwartssignal nur dann auslöschen, wenn beide gleich stark waren. Dann durfte sich weder die Helligkeit, noch die Farbe in zugehörigen Retinaquadrat verändern. Es durfte sich dort genau genommen nichts ändern.
Änderte sich jedoch die Farbe in diesem Retinaquadrat, weil ein andersfarbiges Objekt sichtbar wurde, oder änderte sich die Helligkeit, weil ein Objekt mit anderer Helligkeit erschien, so feuerte diese konkrete Outputzelle nicht mehr. Denn in allen visuellen Modulen befand sich plötzlich das Neuron mit der höchsten Feuerrate in einer anderen Höhe, falls sich die Farbe oder die Helligkeit änderte. Der neue Output musste daher schwächer sein.
Wenn im zugehörigen Retinaquadrat eine geneigte Gerade auf die Netzhaut projiziert wurde, und diese ihren Anstellwinkel oder ihren Nullpunktanstand (zur Quadratmitte) veränderte, so wanderte der Ort der maximalen Erregung von der aktuellen Orientierungssäule zu einer anderen, die nunmehr am stärksten feuerte. Die Feuerrate des aktuellen Neurons wurde schwächer, das Gegenwartssignal war schwächer als das zeitverzögerte Vergangenheitssignal. Hier war das Restsignal gleich null.
Aber es gab ein neues Outputsignal, denn nun feuerte die Orientierungssäule, die zuvor stumm war. Ihr Vergangenheitssignal war das Nullsignal und konnte das neue Gegenwartssignal nicht auslöschen. Genau diese neue Orientierungssäule feuerte nun ungehemmt ihren Output in die sekundären Gebiete des Cortex.
In demjenigen sekundären visuellen Cortex, der den visuellen Output des ventralen Thalamus empfängt, zeigen stark feuernde Neuronen also Bewegungen und Veränderungen an. Doch diese Neuronen haben unterschiedliche visuelle Submodalitäten: Hell/Dunkel, Blau/Grün, Grün/Rot und Rot/Blau.
Verschiedene Modalitäten haben jedoch die Tendenz, sich im Verlauf der Evolution aufzutrennen, aufzuspalten, sich neu zu ordnen. Daher gibt es in demjenigen sekundären Cortex, der Bewegungen analysiert, ebenfalls eine Aufspaltung nach diesen Submodalitäten.
Bedenken sollte man insbesondere, dass (nach meiner Ansicht) das Dickenwachstum der Cortexrinde vor der Ausdehnung in die Breite erfolgte. Es könnte sogar sein, dass bei Reptilien gar kein Breitenwachstum erfolgte und bei Vögeln nur ein sehr geringes. So wären deren Cortexgebiete vorwiegend Divergenzschichten mit vertikaler Signalausbreitung. Dies habe ich bereits in meiner vorigen Monografie als Hypothese formuliert.
Wenn die Breitenausdehnung evolutionär viel später erfolgte als das Dickenwachstum, würde man die dadurch hinzugewonnenen oder neu gebildeten Neuronen durchaus als eigene Modalität betrachten dürfen. Dann wären die Resultate der vertikalen Signaldivergenz eine eigene, ältere Modalität (Blobs), und die Resultate des Breitenwachstums, also die Orientierungssäulen oder Interblobs wären auch eine selbständige, jedoch jüngere Modalität. Diese zwei verschiedenen Modalitäten würden sich im sekundären Cortex auftrennen und eigenständige Gebiete in Anspruch nehmen. Dies scheint tatsächlich der Fall zu sein, wie nachfolgende Überlegungen zeigen.
Die Mittelneuronen eines jeden quaderförmigen Analysemoduls, durch die Werte x = 0 und y = 0 gekennzeichnet, projizieren in zwei Gebiete: Eines repräsentiert die Helligkeit eines Objekts (wenn dieses die gesamten vier magnocellularen retinalen Ganglienzellen komplett ausfüllt) und eines für die Farbe (wenn dieses Objekt ebenfalls die vier parvocellularen retinalen Ganglienzellen komplett ausfüllt). Da es zu jedem Retinapunkt nur wenige Helligkeiten und Farben dieser Erkennungsart gibt, bilden diese Neuronen einen schmalen langen Streifen. Die Retina wird rechteckig auseinandergezogen. Anstelle eines Bildpunktes gibt es eine Anzahl Helligkeitsneuronen und ebenso von Farbneuronen nebeneinander. Die Retina wird zum rechteckigen Abbild mit relativ geringer Höhe. Besitzt die Retina zum Beispiel 500 Bildpunkte in der Höhe und 500 in der Breite, so gibt es in diesem Gebiet beispielsweise ein Rechteck mit 500 Neuronen in der Breite und 500 * 20 = 10000 Helligkeitsneuronen in der Länge. Ein farbempfindliches Gebiet benötigt für die sieben Regenbogenfarben dann ein Rechteck aus 500 Neuronen in der Breite und 3500 in der Länge. Um 500 Neuronen in der Breite unterzubringen, benötigt man für die recht kleinen Cortexneuronen nicht viel Platz. Daher finden wir dort schmale Streifen, die auf Veränderungen der Helligkeit und der Farbe reagieren.
Anders ist es mit dem Output derjenigen Orientierungssäulen, die auf Linienelemente reagieren. Hier wird der Output den Basalganglien übergeben und kommt zeitverzögert zum ventralen Thalamus zurück, wo diese die Vergangenheitssignale die Gegenwartssignale hemmen. Damit zeigen diese Überlagerungsneuronen eine Veränderung der Linienelemente sowohl im Winkel als auch in Helligkeit und Farbe an.
Wenn nun die Retina wiederum beispielhaft aus 500 mal 500 Bildpunkten besteht, gibt es 250000 Bildpunkte insgesamt. Wenn es zu jedem Bildpunkt 100 Orientierungssäulen gibt, können 100 verschiedene Winkel erkannt werden. Insgesamt also 25000000, also eine 25 Millionen
Nun gibt es zu jeder dieser Orientierungssäulen je vier Submodalitäten: Hell/Dunkel, Blau/Grün, Grün/Rot und Rot/Blau. Das sind schon 100 Millionen Varianten. Dies ist noch zu multiplizieren mit der Anzahl der Helligkeitsstufen und der Farbvarianten. Diese Zahl ist riesig.
Daher gibt es wenigstens einen breiten Streifen von Neuronen, die die Hell-Dunkel-Bewegung von Konturen (farbneutralen Richtungselementen) anzeigen. Ebenfalls gibt es einen noch viel breiteren Streifen, der die Farbe von Konturenbewegungen signalisiert. Wenn sich diese Neuronen in der einen Richtung so anordnen, wie sie im Farbmodul in der Vertikalen angeordnet sind, ist dieser „Farbstreifen“ wesentlich breiter als der Hell-Dunkel-Streifen.
Insgesamt gibt es drei verschieden breite Streifen, die ihren Input teils über den Umweg der Basalganglien erhalten und die bewegungssensitiv sind.
Bei den Orientierungssäulen ist die Bewegungsmeldung per Aktionspotentialfolgen am stärksten, wenn ein Linienelement parallel in die Richtung verschoben wird, die mit dem Linienelement einen rechten Winkel bildet. Bewegt sich das Linienelement jedoch entlang der Linie, die es selbst bildet, reagiert die zugeordnete Orientierungssäule jedoch auf zwei verschiedene mögliche Arten:
- Die Linie ist so lang, dass sie die beteiligten Ganglienzellen noch genauso bedeckt, wie sie es vor ihrer Bewegung tat. Dann feuert die „Bewegungs-Orientierungssäule“ nicht: Es hat sich nichts geändert.
- Reicht die Linienlänge nicht aus, um nach der Bewegung die Ganglienzellen zu bedecken, so fällt plötzlich die Abdeckung weg. Das Neuron feuert stark und zeigt uns damit an: Linienende erreicht, Linie verlässt die Ganglienzelle, Änderung erkannt.
Die letztere Eigenschaft nennt man Endinhibition. Alle Neuronen, die Input von den Basalganglien erhalten und im breiten Streifen der richtungsempfindlichen Neuronen liegen, reagieren, wenn durch Bewegung einer Linie in ihre eigene Linienrichtung das Ende der Linie erreicht wird und dadurch die Verdeckung der Ganglienzellen wegfällt.
Die Basalganglien ermöglichen spezifische Bewegungs- und Richtungserkennung in allen Modalitäten. Ihr Output erreicht ausgewählte Gebiete im Thalamus, wo er mit einer Kopie der Originalsignale überlagert wird. Der Output der Differenzabbildung erreicht sekundäre Gebiete, die der Bewegungs- und Richtungsanalyse von Objekten beliebiger Modalitäten dient.
Da im Fall der Orientierungssäulen nicht nur die Veränderung des Winkels von Linienelementen erkannt wird, sondern es sogar eine besonders bevorzugte Bewegungsrichtung während der Analyse gibt, kann das Gehirn aus dieser Information auch die Ortsveränderung ermitteln. Es ermöglicht die Wahrnehmung, dass sich etwas bewegt hat. Es kann jedoch auch die Bewegungsrichtung ermitteln. Doch dazu muss es in der Lage sein, die während der Signaländerung ermittelte Bewegungsrichtung zu speichern und später wieder zur Verfügung zu stellen. Diese Signalspeicherung erfolgt nach meiner Ansicht vorwiegend im Cerebellum, teilweise jedoch auch im Hippocampus.
Die Speicherung im Hippocampus habe ich zwar inhaltlich verstanden, jedoch fehlen mir noch wichtige Informationen, um den Algorithmus verständlich zu beschreiben. Es ist jedoch vorhersehbar, dass es im Hippocampus eine Neuronenorganisation in sechseckigen Formationen geben muss, die der Verschlüsselung von Orten dienen. Diese sechseckigen Neuronenformationen sind ein Tribut an die sechseckigen Schwerpunktmodule, die wiederum eine Folge von sechs Augenmuskeln der Wirbeltiere sind. Beteiligt ist das mehrstufige Mittelwertsystem des Wirbeltiergehirns. Verschieden große Sechseckpopulationen im Hippocampus entsprechen wahrscheinlich unterschiedlichen Stufen des Mittelwertsystems, also unterschiedlichen „Vergrößerungsstufen“.
Diese Verschlüsselung ist unabhängig von der aktuellen Orientierung um Raum. Die hippocampale „Landkarte“ ist quasi unbeweglich im Gehirn verankert, nur der eigene Aufenthaltsort wird in dieser Karte durch feuernde Neuronen gekennzeichnet und wandert daher bei Eigenbewegungen hin und her. Vielleicht finden ja manche Leser Gefallen daran, die Grundlage der Verschlüsselung von Raumkoordinaten im Hippocampus gemeinsam mit mir zu analysieren. Ein wesentlicher Aspekt hierbei ist das hippocampale Lernen, also die Prägung über LTP und LTD, die ähnlich verläuft wie im Pontocerebellum. Auch hier sind tetanische Mittelwertsignale der neuronale Schreibbefehl im neuronalen Netz.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan