Gehirntheorie des Menschen
ISBN 978-3-00-068559-0
Monografie von Dr. rer. nat. Andreas Heinrich Malczan
6 Konvergenzmodule - inverse Divergenzmodule
Für dieses Kapitel müssen wir unsere Einsichten über die Entwicklung des Gehirns aus dem frühen doppelten Strickleitersystem der segmentierten Bilateria aktivieren. Dort stiegen die sensorischen Signale kopfwärts und bogen in jeder Etage zusätzlich zur Seite zum motorischen Teil. Auch im ersten Segment, der zum Cortex wurde, war dies so.
Jedes sensorische Signal musste von der sensorischen Cortexseite auf die motorische Cortexseite ziehen. Und da der Cortex inzwischen in Modalitätenlappen und Modalitätenstreifen in jedem Gyrus zerfallen war, stand jede Modalität vor dieser Aufgabe. Sie musste zur motorischen Seite wechseln, damit ihre Signale die zugehörigen Motoneuronen ansteuerten.
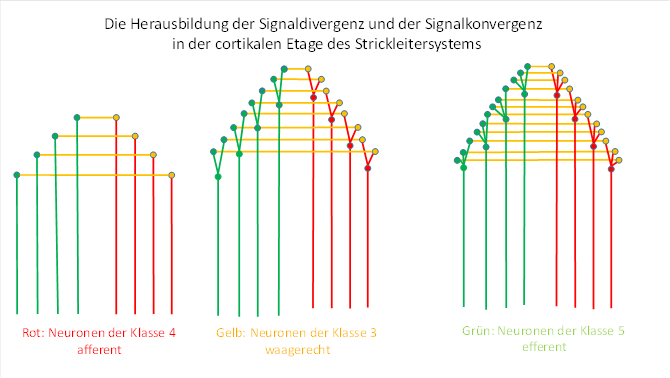
Abbildung 38: Divergenz und Konvergenz in der cortikalen Etage
So konnte jede Modalität mit den Signalen ihrer Rezeptoren beispielsweise eine Bewegung zur Beute bewirkten, aber auch Flucht vor Fressfeinden.
Mit der Entstehung der Signaldivergenz auf der sensorischen Seite der cortikalen Etage des frühen Strickleitersystems wurden die aufsteigenden Signale divergent auf die Neuronen der Klasse 3 verteilt. Dies vollzog sich sicherlich auch schrittweise. Anfangs versorgte ein Neuron der Klasse 4 genau ein Neuron der Klasse 3, dessen Axon zur motorischen Seite zog und an genau einem Neuron der Klasse 5 endete. Später verteilte sich das aufsteigenden Signal auf 2 Neuronen, die zur motorischen Seite zogen. Sie hatten auf der motorischen Seite jedoch nur ein Zielneuron, an dem sie beide gemeinsam andockten. Als ein drittes Neuron der Klasse 3 mit einbezogen wurde, trafen drei Axone auf der motorischen Seite ein und kontaktierten das gemeinsame Zielneuron. Dies ist in der obigen Abbildung dargestellt.
Anfangs diente die Signaldivergenz sicherlich ausschließlich der Erhöhung der Ausfallsicherheit. Erst später – bedingt durch den Abstand zwischen den Input- und Outputneuronen – trat als Folge der nichtlinearen Signaldämpfung eine Extremwertcodierung des Outputs auf, die den Tieren zum Vorteil gereichte.
Je weiter diese Entwicklung voranschritt, umso größer wurde die Anzahl der Neuronen der Klasse 3. Während auf der sensorischen Seite durch die Signaldivergenz die verschiedenen Divergenzmodule entstanden und eine viel feinere Analyse der Signale ermöglichten, musste auf der motorischen Seite eine entsprechende Signalkonvergenz entwickelt werden. So entstanden auf der motorischen Seite die Konvergenzmodule. Sie machten die sensorische Signaldivergenz rückgängig.
Auf der motorischen Seite fand eine Signalkonvergenz statt, welche die Signaldivergenz der sensorischen Seite komplett rückgängig machte. So erhielt das angeschlossene Motoneuron, egal, in welchem Segment und auf welcher Körperseite es sich befand, den Input, den es vor der Ausbildung der Signaldivergenz erhalten hatte. Die motorischen Reaktionen auf das sensorische Signal blieben erhalten.
Wir haben festgestellt, dass durch die Signaldivergenz neue Submodalitäten entstanden: Helligkeit, Farbe, Orientierungswinkel von Linienelementen. Bewegung usw. Die ursprüngliche Modalität zerfiel dadurch in neue Submodaltäten.
Wann immer eine Modalität durch Signaldivergenz in Divergenzmodulen in neue Submodalitäten zerfiel, wurden auch die neuen Signale zum motorischen Teil des zugeordneten Cortex gesendet. Es war genau die Cortexregion, zu der die ursprünglichen Ursignale durch die Axone der Outputneuronen geschickt wurden.
Fand nun in der Schicht der Sensoriksignale eine Signaldivergenz statt, so zogen die divergierten Signale genau in dieses Gebiet und konvergierten wieder auf die ursprünglichen Outputneuronen, die zuvor genau die Ursignale erhalten hatten.
Im Fall der Signaldivergenz von motorisch wirksamen Signalen zogen diese von der sensorischen Seite – wo sie etwa als Muskelspannungssignale eintrafen – zur motorischen Seite und konvergierten auf die Beetz-Zellen des motorischen Cortex. Deren große Dendritenbäume empfingen die zuvor divergierten Signale und fassten sie wieder zusammen. So entstand das Ursignal auf der motorischen Seite durch eine starke Signalkonvergenz. Diese machte die Signaldivergenz wieder rückgängig.
Wenn nun ein Divergenzmodul zu einem Dickenwachstum oder zu einem Flächenwachstum oder (meist) zu beidem führte, so fand genau diese Veränderung nicht nur auf der sensorischen Seite statt, sondern in gleichem Umfang auf der motorischen Seite. In den Zielgebieten eines Moduls trafen die Signale genauso auf ein analog aufgebautes motorisches Modul, welches die Signaldivergenz rückgängig machte.
Empfing beispielsweise ein würfelförmiges Farbmodul Rot/Grün mit räumlicher Signalausbreitung den Retinainput von insgesamt acht Ganglienzellen und erstellte daraus beispielsweise 1000 verschiedene Outputsignale (10 Neuronen in der Breite, 10 in der Länge und 10 in der Höhe – Kapitel 1.1.5), so bestand es aus 10 Schichten, die jeweils übereinander lagen und von denen jede quadratisch aufgebaut war und 10 Neuronen in der Breite und 10 in der Tiefe enthielt.
Diese Neuronen projizierten (unter anderem) zur motorischen Seite, die dann ebenfalls aus 10 Schichten von Neuronen bestand. Dort kontaktierten sie 1000 Neuronen in einem Würfel, der aus 10 Neuronenschichten a 100 Neuronen bestand, die jeweils in einem Quadrat von 10 mal 10 Neuronen angeordnet waren. Doch beide Würfel, der sensorische und der motorische, waren spiegelbildlich zueinander angeordnet. Die Ebene, die senkrecht zwischen den Würfeln verlief, war die Spiegelebene.
Nun konnten die Signale der Neuronen der Klasse 3 zum motorischen Teil ziehen und dort die 1000 Interneuronen in der Schicht 5 kontaktieren, ohne dass sich ihre Axone in den Weg kamen und Unordnung entstand. Sie zogen auf der sensorischen Seite einfach nach unten, bis sie die Cortexschicht verlassen hatten. Danach zogen sie auf halbkreisähnlichen Bahnen zur Gegenseite nach oben ins motorische Gebiet. Von dort ging es senkrecht weiter, bis jedes Axon die Höhe erreicht hatte, in der sich auch das betreffende Zielneuron der Schicht 5 befand.
Die Axone der sensorischen Neuronen der Klasse 3 erregten diese Interneuronen, deren Axone die Signale zu den Würfelecken übertrugen. So gab es im Konvergenzmodul wieder die acht Signale, die zuvor das Farbmodul gespeist hatten. Sie konnten von den Outputneuronen der Klasse 5, die sich in den acht Würfelecken befanden, empfangen werden und wurden nun den ursprünglichen Signalempfängern – den Motoneuronen – zugestellt.
Diese Konvergenzmodule – von mir auch als inverse Divergenzmodule bezeichnet, erzeugten aus den Signalen der neuen Modalitäten wieder die Signale der alten Modalitäten. Diese Behauptung muss jedoch mathematisch bewiesen werden.
Hier sollten wir die These verfolgen, dass die Schichtung des Cortex erhalten blieb. Nur dadurch konnte ein inverses Modul die Arbeit des vorherigen Analysemoduls rückgängig machen.
Natürlich hätten die Ursignale auch direkt zur motorischen Seite ziehen können. Da aber die Ursignale aus Rezeptoren kamen, hätten stattdessen auch die Rezeptoren direkt die Muskeln steuern können. Ein Gehirn wäre dann gar nicht nötig gewesen. Jeder Rezeptor hätte – wie in Urzeiten – einfach ein Motoneuron ansteuern können.
Daher muss die Signalanalyse in den cortikalen Divergenzmodulen einen höheren Nutzen gehabt haben, sonst wäre sie nicht entstanden.
Der erste Nutzen entstand zur Urzeit, als Lebewesen die Fähigkeit erlangten, viele Farben zu erkennen. Dies erleichterte die Erkennung von Futter. Während Pflanzenfresser von grünempfindlichen Sehpigmenten ihren Nutzen zogen, weil das grüne Chlorophyll Pflanzen verriet, konnten Fleischfresser die rote Farbe des Blutes sicher detektieren. Als einige Spezies sowohl die Farbe Grün als auch die Farbe Rot erkennen konnten und auch ihre Zwischenfarben erkannten, waren Allesfresser im Vorteil. Ob diese simple Betrachtungsweise einer kritischen Analyse standhält, ist fraglich. Doch Hypothesenbildung ist das Geschäft der Forscher. Jede Hypothese kann – falls sie sich nicht bewährt – über Bord geworfen werden.
5.1 Das Konvergenzmodul für das Farbmodul mit vertikaler Signalmischung
Wir untersuchen nun die Funktionsweise eines Konvergenzmoduls mit vertikaler Signalmischung, kurz auch als vertikales Konvergenzmodul bezeichnet. Wir können es ebenso auch als inverses Divergenzmodul bezeichnen, vor allem deshalb, weil darin auf seine Funktionsweise Bezug genommen wird. Sein Input entstammt einem Divergenzmodul. Über Axone der Neuronen der Klasse 3 schickt dieses Modul seinen Output zur motorischen Seite in das Konvergenzmodul. Sowohl im Divergenzmodul aus auch im Konvergenzmodul breitet sich die Erregung (vornehmlich) in vertikaler Richtung aus. Der Grund ist die Signalverwandtschaft der Inputsignale eines Retinapixels.
Die Retina wird in die x-y-Ebene abgebildet. Jedem Retinapunkt PR(x,y) werden im Modul zwei Punkte zugeordnet. Das Rot-On-Signal mit der Feuerrate f1 erreicht das Modul im Punkt PM(x,y,1) auf der Höhe z = 1. Das zugehörige Grün-On-Signal trifft im Modulpunkt PM(x,y,0) ein, also auf der Höhe z = 0. Da beide Signal signalverwandt sind, empfangen alle Neuronen mit den Koordinaten y und x diese zwei Inputerregungen, egal auf welcher Höhe z sie sich befinden. Durch Überlagerung beider Erregungen entsteht in der vertikalen Neuronensäule ein Erregungsmaximum, dessen Höhe die Farbe, also auch die (mittlere) Frequenz im Rot-Grün-Bereich verschlüsselt. Grund ist die nichtlineare Übertragungsfunktion, die zudem streng konkav ist. Dies garantiert ein globales Erregungsmaximum.
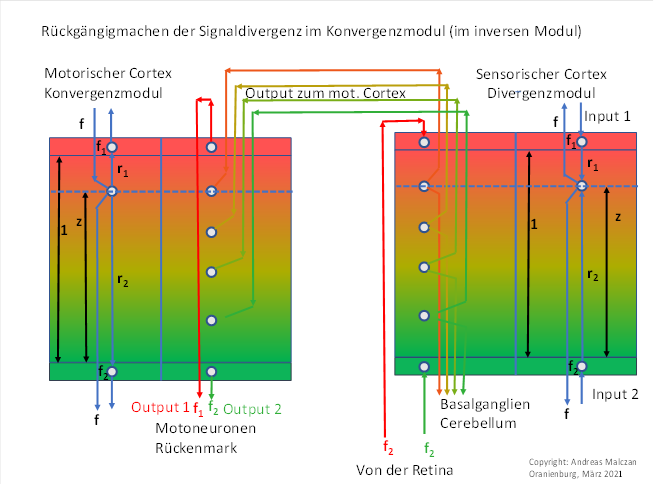
Abbildung 39: Konvergenzmodul für das Farbmodul Rot-Grün mit vertikaler Signalausbreitung
Jedes der zwei Module der obigen Abbildungen ist zweigeteilt. Außen ist das Prinzipschaltbild für drei beteiligte Neuronen dargestellt, die Größen h, r1 und r2 sind durch schwarze Pfeile erklärt und die Dicke der Outputschicht mit 1 gekennzeichnet.
Wir wählen als Beispiel das Farbmodul mit vertikaler Ausbreitung, also ohne Orientierungssäulen. Es dient der reinen Farberkennung. Sein Output zieht über die Axone der Neuronen der Klasse 3 zur motorischen Seite in das zugehörige Konvergenzmodul. Alle Farbsignale einer Augendominanzsäule ziehen wohlgeordnet zur motorischen Seite und kontaktieren (vermutlich) in der Schicht 5 – der Inputschicht – wieder je ein Outputneuron pro Farbsignal. Dabei ordnen sich die zu einer Mischfarbe gehörenden Inputneuronen genau in der Reihenfolge von oben nach unten an, wie sie auch im sensorischen Divergenzmodul angeordnet waren. Die Farbschichtung bleibt also erhalten.
Wir wissen, dass der Output des Divergenzmoduls maximumcodiert ist. Nur ein Neuron in der gesamten vertikalen Säule ist maximal erregt. Es hemmt über die Nachbarhemmung die übrigen Outputneuronen der Outputsäule. So ist von den vielen Farbleitungen zum motorischen Cortex immer nur eine aktiv.
Diese Farbleitungen - also die Axone der Neuronen der Klasse 3 des Divergenzmoduls – verlaufen in Abweichung zur obigen Skizze unterhalb der Cortexschicht auf langen, gut mit Myelin isolierten Axonen zum motorischen Cortex. Hierbei bilden diese Axone in etwa halbkreisförmige Verbindungen, wobei die untere Hälfte eines Halbkreises die Bahn der Axone beschreibt.
Wir erinnern uns: Nur ein Farbneuron in einer Höhe z über der x-y-Ebene in der Neuronenzsäule des Divergenzmoduls ist maximal erregt. Nur sein Output erreicht die motorische Seite, die übrigen Farbsignale werden ein Opfer der Nachbarhemmung.
Im Konvergenzmodul trifft dieses Farbsignal auf ein Inputneuron, welches ebenfalls die Höhe z von der x-y-Ebene hat, und erregt es mit der Feuerrate f. Seine Erregung breitet sich nun im Modul nach oben und nach unten aus. An der oberen Begrenzungsfläche der Inputschicht liegt eine dünne, einlagige Neuronenschicht von Outputneuronen, an der unteren Begrenzungsfläche ebenfalls. Dort befinden sich die beiden Outputneuronen des Moduls, während die Inputneuronen wieder wie eine Perlenschnur von oben nach unten verlaufen.
Während der Ausbreitung wirkt die abstandsabhängige Dämpfung. Das obere Outputneuron – es steht für die Farbe Rot – hat den Abstand r1 vom Inputneuron. Wir haben die Dicke der Inputschicht auf den Wert 1 normiert (eine andere Schichtdicke würde an der prinzipiellen Funktionsweise nichts ändern). Das obere Outputneuron hat zum Inputneuron den Abstand r1, das untere den Abstand r2. Die Inputfeuerrate hat den Wert fD.
Damit gilt für die Feuerraten f1 und f2 der beiden Outputneuronen die Gleichung
![]()
![]()
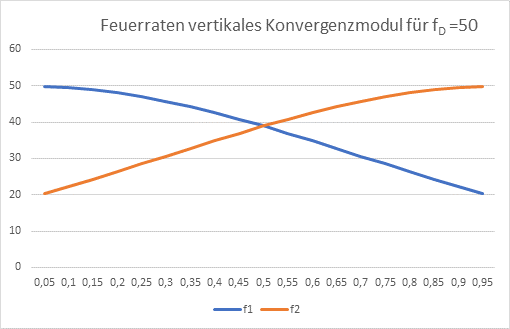
Abbildung 40: Feuerraten vertikales Konvergenzmodul
Im Divergenzmodul ändert sich die Höhe z des Outputneurons mit der maximalen Feuerrate in Abhängigkeit von den zwei Inputfeuerraten f1D und f2D. Dies ist in der nachfolgenden Abbildung dargestellt.
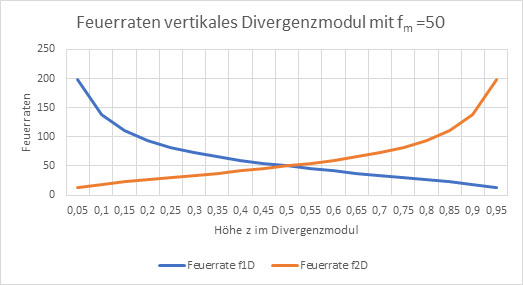
Abbildung 41: Feuerraten vertikales Divergenzmodul
Im Vergleich erkennt man, dass das Konvergenzmodul die ursprünglichen Feuerraten, die als Input des Divergenzmoduls auftreten, nicht exakt, sondern nur angenähert wiederherstellt. Dieser Fehler ist jedoch gewiss viel günstiger als die Alternative, bei der gar kein Output von der sensorischen zur motorischen Seite ziehen würde. Insofern war die Entwicklung von Konvergenzmodulen wichtig für die weiteren, meist motorischen Reaktionen des Tieres auf sensorischen Input.
Doch erst durch die Entwicklung von Speichermodulen brachte die Feinanalyse aller Modalitäten und die Erzeugung neuer Submodalitäten durch Divergenzmodul echte Vorteile. So konnte allein die Vorstellung einer Beute – d. h. die Aktivierung eines gespeicherten Abbildes der Beute – genau die Reaktionen auslösen, die eine reale Beute bewirkte. Beim Menschen können auf diese Weise Abbildungen von gespeicherten Zuständen und Signalkombinationen zum Cortex gesendet werden und dort Aktivitäten aller Art auslösen – vom Träumen bis zur Suche nach Nahrung, Wasser oder gar Sex.
Daher wird im Kapitel 7 das Speichermodul des Pontocerebellums behandelt. Dort werden große Teile aus meiner Monografie „Gehirntheorie der Wirbeltiere“ fast wörtlich übernommen. Eine Ausweisung als Eigenzitate unterlasse ich, da die Urheberrechte an diesen Texten bei mir liegen.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan