Human brain theory
ISBN 978-3-00-068559-0
Monograph of Dr. rer. nat. Andreas Heinrich Malczan
1 The evolutionary history of the brain - the concept of segments and neuron classes
The vertebrate brain probably developed from a centralised nervous system of the simplest multicellular organisms. In these, cell differentiation was already far advanced and there were already many cell types with a wide variety of functions.
In the course of evolution, the number of body cells increased steadily, so that the number of cells was many times greater than the number of different cell types. Here it proved expedient to group cells with the same task together spatially, so that organ formation manifested itself.
Nerve cells had also specialised. One group provided input to the existing life support systems. Throughout the body, sensors measured important body variables and translated them into firing rates. For example, oxygen saturation was monitored and control signals for the heart's pumping rate were derived from this. Many other variables required to maintain vitality were also monitored.
The associated receptors sent their signals to the control centres, which we interpret as mean value centres. In them, the signals from the entire body were combined, i.e. a kind of mean value was formed, which in turn represented the control signal and triggered corresponding reactions. In these mean value centres, there were mean value neurons that combined and integrated the incoming signals from the different parts of the body. We still find such mean value centres in the human brain today, for example in the formatio reticularis.
We assume that the first simple multicellular organisms, whose lineages led to the vertebrates, among others, were already bilaterally symmetrical. Therefore, each of the different mean centres was present on both halves of the body. Bilaterality was also preserved in the vertebrates.
These simple multicellular organisms were (among other things) the precursors of future vertebrates. Animals - in contrast to plants - could move actively. We therefore assume the existence of muscles that made movement possible. The muscles, for their part, needed control signals for their activation. The control signals were provided by receptors. The neurons for receiving the sensory control signals formed a separate nucleus on each side of the body, which can be interpreted as a neural organ and which we call the sensory centre. Its neurons received input from the receptors that could cause motor actions. These were, for example, tactile signals, olfactory signals, visual signals, but also signals from receptors within muscles that measured their contraction.
The output of the sensory centre of one half of the body was delivered to the motor neurons that supplied contraction signals to the muscles of the same side. These neurons also began to concentrate spatially and formed their own neuron nucleus, which we call the motor centre.
Within the sensory and motor centres, mean value neurons developed, which took up the neuronal excitation located in the centre with a larger dendrite tree and transferred it to the mean value centres. We interpret these neurons as mean neurons, recognisable by the larger dendrite tree. It was convenient to transmit the integrated sensory and motor excitation to the mean centres of the same half of the body. In this way, the activity of the mean centres could be adapted to the sensory and motor activity of the animal.
Thus, these multicellular organisms, which led to the vertebrates, had several midline centres on each half of the body, one sensory and one motor centre each.
They had several classes of projection neurons, which we number. Since we later find these classes of neurons in the cortex of the human brain (and in the brains of all vertebrates), we give them the class numbers corresponding to the cortex layer in which we find these projection neurons.
We assign class number 6 to the mean neurons in the sensory and motor centres. We find them in the sixth layer of the cortex, where they form large pyramidal cells, absorb the input from the environment and transmit it to the midline structures of the brain. Evolutionarily, they are the oldest class of neurons, so they form the lowest, inner layer in the cortex.
We assign class number 3 to the projection neurons of the sensory centres. We find them in the third cortex layer of humans, where they receive the ascending sensory input from the trunk and head receptors and transmit it to the motor side of the cortex. Likewise, we find them in all segments (vertebrates are segmented), where they receive the sensory input that reaches them via the sensory ganglia.
We assign the class number 5 to the projection neurons of the motor centres. We find them in the human cortex on the motor side in the fifth layer. They receive input from the sensory neurons of class 3, which send their axons to them. Furthermore, we find them in all segments, there they receive the output of the sensory neurons and in turn control the motor neurons.
In addition to the projection neurons, there may already have been interneurons at that time, which enabled interactions between projection neurons. They were always located within a neuronal nucleus. The inhibitory interneurons could enable lateral inhibition. Each projection neuron could activate inhibitory interneurons in its immediate vicinity, which in turn inhibited neighbouring projection neurons. Thus, neighbouring projection neurons were in neuronal competition with each other and inhibited each other - with the interposition of the inhibitory interneurons. In this way, stronger signals prevailed over weaker ones.
In this monograph, we also assume an exchange of signals between the two halves of the body. This was realised by class 2 projection neurons. We find them in the cortex in the second layer, where they (often) move with their axons to the contralateral half of the cortex (or to neighbouring areas) and deliver their signals. They are excitatory. Similarly, we find them in the segments of the spinal cord, where they provide input to the contralateral side of the segment, enabling motor reflexes, for example.
The two halves of the body were in neuronal competition with each other via the projection neurons of class 2. Each neuronal nucleus projected excitatory via them into the same nucleus of the opposite side. Thus, the sensory nucleus projected excitatory through class 2 neurons into the sensory nucleus of the opposite side. The neuronal competition arose because these signals ended at inhibitory interneurons, which in turn caused lateral inhibition. The same was true for the motor nucleus of each side of the body and for the different mean nuclei. The left and right sides of the body were in neuronal competition with each other.
That made sense. Stronger signals from one half of the body were preferred and suppressed weaker signals from the opposite side. This is how selection took place.
Since the growth of the axons that realised the neuronal connections was controlled by marker substances, each half of the body had to have its own marker in order for the projection neurons of one half of the body to find the target structures in that half of the body. Therefore, a class 2 projection axon could not simply move from one side of the body to the contralateral side. It could only move to the parting plane that separated the two sides of the body. There, another class 2 projection neuron had to accept the transmitted excitation. However, because this neuron was already on the opposite side, albeit close to the separation plane, its axon could find the target structure.
The signal transfer from one side of the body to the other for the purpose of contralateral inhibition thus always took place via intermediate projection neurons that were located near the separation plane of the two halves of the body. These neurons also each formed a nucleus, which is called a side-switch nucleus. We find such side-switch nuclei again in the human brain. The nucleus olivaris receives the output of the nucleus ruber and transmits it to the contralateral cerebellum, so it is a typical side-switch nucleus. The side-switching nuclei of the two halves of the body were located closely adjacent to each other at the dividing plane of the two halves. In the course of later evolution, they could also merge with each other, so that the signals of both halves of the body were available there simultaneously and could interact with each other.
Another class of neurons may also have existed at that time. The median nuclei monitored and controlled the vital functions. If the metabolism caused a lack of the substances that served as energy suppliers, a supply of food was necessary. It was favourable if the relevant mean nucleus not only detected the lack of energy, but at the same time stimulated the sensory and motor functions. An excitatory back-projection of the mean nucleus into the sensory and motor centre of the respective side of the body led to the pre-excitation of the neurons. This was able to lower the response threshold, and stimuli were now responded to earlier. Possibly, mean nuclei could even evoke action potentials in the sensory or motor nuclei and activate the body directly. This is how hunger led to the search for prey.
The action of the mean nuclei on the sensory and motor nuclei took place via class 1 neurons. These neurons were located on each side of the body in the sensory and motor nuclei. They received the mean excitation from the mean centres on the same side of the body and passed it on to the projection neurons of the sensory and motor nuclei, causing them to be activated. We find these activation neurons in the human cortex in the first layer, where they can activate the surrounding neurons with widely branching dendrites. In the course of evolution, they could have arisen as the last neuron class, which is why we find them in the cortex in the uppermost, first layer. The development of these activation neurons made it possible to use internal system states to activate sensory and motor functions, which was an enormous advance.
Thus, at a sufficiently high stage of development, the simple multicellular organisms already possessed four of the six neuron classes of the human brain. Neuron classes 1, 2, 3 and 6 existed at that time. How did the remaining two neuron classes come into being?
I believe that this occurred during the transition from multicellular to segmented animals.
We assume that in the Bilateria, whose lineages led to the vertebrates, there was a generational change during reproduction. Sexual reproduction constantly alternated with asexual reproduction. In asexual reproduction, we imagine that a kind of bud formed at the end of the animal, from which an offspring emerged that later separated from the mother animal. If the separation failed to take place, a double being arose, the smallest colony of two identical beings.
In my monograph "Brain Theory of Vertebrates" I describe this in chapter 2.5 as follows:
"The asexual reproduction of these living beings thus involved the formation of a bud from the budding zone, which ultimately developed into a complete second living being. The final step was the separation of this second living being so that it became an offspring.
This separation was an important step that was anchored in the genetic material and had to be controlled biologically somehow. If the coding of the individual steps in the DNA was linear in the order in which they occurred, the code for the separation was again at the end of the coding chain. It is possible that sometimes this part was lost during the replication of the DNA or was skipped due to special causes. Then a disturbance occurred here, and so a double being arose instead of an offspring. If the unseparated daughter creature was also viable afterwards, because it was able to take in food (or was also supplied) and had all the necessary vital functions, this reproductive error was not particularly dramatic. Practically, a (very small) colony of two identical living beings was created, which remained connected to each other. If in the course of evolution such strangulation defects occurred repeatedly and accumulated, they could manifest themselves hereditarily. Then the last step of the asexual reproduction process, the strangulation of the successor emerging from the bud, was genetically deleted from the hereditary programme and the emergence of two-segmenters manifested.
One could also speak of a new living being consisting of two segments having emerged here."
It is not difficult to imagine how colonies of three, four and more animals developed in the same way. If this colony formation was included in the inheritance programme and maintained, then from a certain stage of evolution these animals could be regarded as segmented animals. They consisted of individual animals, but at the latest with the development of a division of labour between the segments, true segmented animals emerged. In these, the failure to separate the daughter creatures created by budding was genetically manifested. A segmented creature also developed in the sexual reproduction path. This is how the segmented bilateria evolved.
An exchange of substances and signals took place between the segments of a segmented animal. On the one hand, shared organs or organ systems developed. For example, the shared intestine, which initially extended from the first to the last segment. Nevertheless, most segments were initially almost completely identical. Simple animals, such as worms, still show this original state. But in the course of evolution, there were major changes in the body plan. What reasons might there have been for this?
I think the main reason for this development was the original presence of an elementary, central nervous system already in the multicellular organisms, which later became segmented animals through colony formation. Only those multicellular organisms whose nervous system already had inhibitory interneurons for lateral inhibition and for contralateral inhibition were able to follow an evolutionary path towards chordates. Another main reason is the formation of a neuronal signal exchange between the segments of the segmented animals, which then gave rise to the chordates.
This needs to be explained.
I have already described that in the bilateral multicellular animals, whose lineages led to the vertebrates, there were neuron classes 1, 2, 3 and 6, which we also find in the cortex layers of humans. As neuronal structures, these animals possessed a sensory centre, a motor centre and several, different mean centres on each side of the body. The latter served as input suppliers for the life support systems, but also had an activating effect on the sensory and motor centres.
With the emergence of segmented bilateria, the described structure was found again in each segment, since (almost) all segments were initially identical. Only the last segment in these animals had an additional budding zone for asexual reproduction.
I assume that the neuronal signals of the receptors of a segment were able to overcome the segment boundaries in the course of evolution. This was due to the close relationship of neighbouring segments, which was transferred to the neurons. Even today, it is observed that the receptor signals of segmented organisms can cross the segment boundaries and end up in neighbouring segments. An example of this are the dermatomes of humans. A dermatome represents the area of skin that is autonomously supplied by the sensitive fibres of a spinal nerve root. The dermatomes also cross the segment boundaries so that they overlap. Here, the information from one segment also reaches the neighbouring segments.
In the course of evolution, special neuron classes developed for the exchange of signals between the segments. One neuron class took over the signal transport in the direction of the head, i.e. the afferent direction. We assign class number 4 to this neuron class. We find them in the human cortex in the fourth layer. There, however, their neurons have become interneurons because the cortex corresponds to the highest head-side segment and thus there is no higher segment to which these class 4 neurons could pass on their excitation.
The exchange of signals in the direction of the tail is carried out by projection neurons, to which we assign neuron class 5. We find them in the fifth cortex layer of humans. There they take over the cortex excitation and project tailwards to the motor neurons.
This reveals a division of tasks between the neurons of class 4 and 5. The neurons of class 4 are sensory projection neurons. They transmit the signals from the sensory centre to the sensory centres of the superior segments.
The neurons of class 5 are motor neurons. They transmit the signals from the motor centres tailwards to the motor neurons of those segments to which the respective motor signals are assigned.
One reason for splitting the vertical signal transmission into two independent types was the control of the growth direction of the axons by markers. Class 4 axons had the opposite growth direction of class 5 axons, presumably responding to a head marker while the others followed the tail marker.
And because the body of these animals is bilaterally symmetrical, the axons of the sensory neurons of class 4 form a nerve cord on each half of the body, running from the tail segment to the head segment and forming a ganglion in each segment, which contains the sensory centre of the segment.
Similarly, the axons of the class 5 motor neurons on each side of the body form a nerve cord that runs from the head segment to the tail segment, forming a ganglion in each segment that contains the motor centre of the segment.
Since in each segment - as in the multicellular organism described above - the sensory centre is linked to the motor centre by the axons of class 3 neurons, the nervous system of these bilateria looks like a rope ladder on each side of the body. This is why it is also called the rope ladder nervous system in the German-speaking world.
Both cord ladders of the two halves of the body are additionally linked by the axons of class 2 neurons, which connect the left and right motor and sensory centres respectively and terminate at inhibitory interneurons to realise contralateral inhibition. When transitioning from one side of the body to the other, one lateral switch nucleus is required per segment and side of the body. A total of four nerve cords are involved, two sensory and two motor. Thus, the classic tetraneural nervous system is present here.
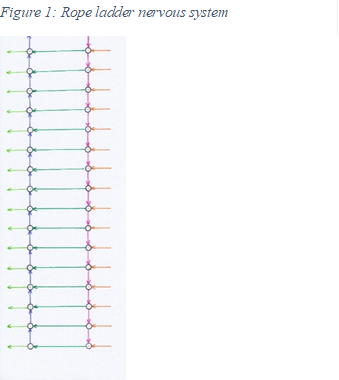 |
We should mention here that in each segment there are the midline systems that receive sensory and motor input and project it in an activating manner into class 1 neurons that are present in both the motor and sensory centres. Here, too, an exchange of signals developed between the segments. For simplicity, we assume that the neurons of classes 4 and 5 were also used for this.
In the course of evolution, the body size of animals grew, the number of receptors grew as well and many new types of receptors also developed. This led to a strong increase in the number of signal-processing neurons in the sensory and motor centres as well as in the mean centres. This was also associated with a strong increase in the number of neurons that enabled the exchange of signals between the segments.
The number of axons of the projection neurons of classes 4 and 5 grew so much that initially broad bands of axons managed the head- and tail-side signal transport. These widened and took on a shape that was as worm-like as the body of the animals at that time. This is how the so-called neural tube came into being. Half of it consists of the axons of the headward projecting neurons of class 4, while the other half consists of the axons of the tailward projecting neurons of class 5. While the axons are arranged on the outside, the associated cell bodies of the neurons occupy the inner space and form the grey matter there. This system is called a neural tube because there is a fluid-filled ventricular space inside. The cell nuclei seek the proximity of this ventricular space, possibly because they obtain their nutrients from it and deliver their metabolic products to it.
In the neural tube, but also in the later spinal cord, the sensory receptors of the animal's body end in the sensory half called the posterior horn, while the motor neurons are located in the neural tube half called the anterior horn. Class 3 sensory neurons project to the motor half via their axons, maintaining the respective side of the body. This is shown in the following figure, the class 2 neurons for contralateral inhibition have been omitted for simplicity.
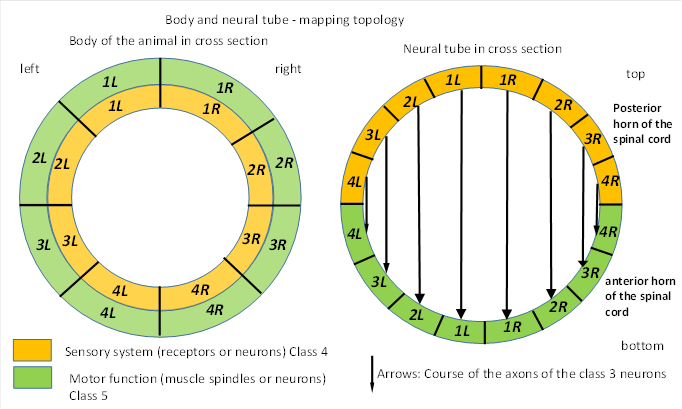
Figure 2: Body and neural tube - mapping topology
Image source: Brain theory of vertebrates, Andreas Malczan, 2020, Fig. 6
The ventricular space leads to the curvature of the axons of neuron classes 2 and 3, as they cannot enter it (Not considered in the above figure).
In the neural tube of those creatures whose lineage led to the vertebrates, in my opinion, a well-ordering of the different neuron classes developed in independent layers from the outside to the inside in the following order:
· Layer 6 of neuron class 6 (mean neurons)
· Layer 5 of the class 5 motor commissural neurons
· Layer 4 of the class 4 sensory commissural neurons
· Layer 3 of the class 3 sensory connective neurons
· Layer 2 of the class 2 motor connective neurons
· Layer 1 of the class 1 activation neurons
Here, layer 1 with class 1 neurons is immediately adjacent to the ventricular space of the neural tube. The neighbourhood of the ventricular space to neuron class 1 seems to be essential for the stratification direction.
It is striking that the neurons in the human cortex are arranged in layers in exactly the opposite order. This is certainly due to the spatial coupling of the class 1 neurons to the outer ventricular system of the cortex.
In every neuronal nucleus, according to our understanding, inhibitory interneurons exist for lateral inhibition. Precisely this fact is an extremely important reason for the specialisation of the segments. Whereas initially almost every organ was found in almost every segment, this is no longer the case in vertebrates, for example. How can this be explained?
This is related to the organisational form of signal forwarding. We divide the input of a segment into own input and foreign input. The intrinsic input comes from the receptors that are present in this segment. Extraneous input comes from the receptors of all the other segments, which are arranged tailwards.
We assume here that the extraneous input in each segment was treated in the same way as the intrinsic input. The intrinsic input terminated at class 3 neurons projecting to the motor centre, but also at class 4 neurons, which pulled towards the sensory centre of the neighbouring segment in the head direction. The extraneous input that reached the sensory centre via ascending axons of class 4 projection neurons was treated in the same way. On the one hand, it terminated at class 3 neurons that projected to the motor centre of the same segment on the same side of the body. On the other hand, it terminated at sensory projection neurons of class 4, which travelled to the neighbouring segment on the head side and terminated there in the sensory centre.
Thus, in those segmented bilateria whose lineages led to the vertebrates, each segment received a copy of all the sensory signals from those segments that were present in the tail direction. The first head segment received the sensory input from all segments of the animal. This is where most of the information was present, this is where the cortex was formed. This will be described in more detail later.
But what happened to the motor signals?
The intrinsic input of a segment arrived at the sensory centre and was passed through to the motor centre. There it met the neurons of class 5, which projected into the motor neurons of this segment. The intrinsic signals had thus reached their targets.
The foreign input came from other, lower-lying segments. The motor targets were located in the origin segment of the signals. Therefore, the foreign input was passed on to the neighbouring segment via downward projecting neurons of class 5. There, the part of the signals that had originally been delivered by the receptors of this segment was passed on to the associated projection neurons of class 5, which passed it on to the motor neurons of the segment. The remaining signals of the extraneous input were passed in descending order from segment to segment via class 5 neurons until they reached their segment of origin, where they could be passed on to the assigned motor neurons.
In the head segment, the motor output was still available for all segments. From segment to segment, this complete output was reduced by the motor signals that had their motor targets in the current segment. The tail segment only received signals that were assigned to its own motor neurons.
This special form of organisation was to prove particularly useful for vertebrates. However, it could be formed in a similar way in other animals.
If the neuronal signals were attenuated during their propagation on the axons of the sensory or motor projection neurons, i.e. if they became weaker with increasing distance, they no longer reached all segments. Then the signal exchange only reached a few neighbouring segments. Such a condition can be observed in worms. This prevented further evolutionary development towards vertebrates.
The sensory signal exchange defined a neighbourhood relationship in which there were subordinate and superior segments. All tail-side segments were subordinate, all head-side segments were superordinate.
In addition to its own sensory input, each segment also possessed the input of all subordinate segments.
Here it is time to take a closer look at this neuronal form of organisation. How were the neurons spatially arranged?
In principle, it is simplest if we imagine that each segment, in addition to its own sensory centre, has a copy of all the sensory centres of the subordinate segments. A collection of sensory centres. Each copy would be constructed in the same way as its original. The head segment would then have as many sensory centres per half of the body as there were segments.
This is how we make the transition from sensory centre to ganglion. A sensory ganglion on one side of the body of a segment consists of the sensory centre of this segment and of the copies of the sensory centres of all subordinate segments. And the cord ladder system consists of such ganglia.
It was the same on the motor side. In the head segment there was the motor centre of the head segment and the copies of all the motor centres of the subordinate segments. They formed the motor ganglion. And in each lower segment there was the motor centre of that segment and the copy of the motor centres of all the subordinate segments.
This rather simple form of organisation was certainly easy to translate into a sequence of commands that provided the building instructions for the nervous system in the animal's DNA.
Now we only have to clarify how the copies of the neuronal centres were arranged in each segment. To do this, we are already imagining a higher state of development and demand the simplest possible form of organisation. Let us consider the right side of the animal's body as an example. Then let the sensory centre be on the extreme right, while to its left the copy of the sensory centre of the neighbouring segment on the tail side is attached. To its left is the copy of the sensory segment of the next, tail-side segment. This is repeated until the copy of the tail segment is attached.
Inside are the copies of the tail-side segments, well-ordered according to ascending segments; outside is the sensory centre of the segment.
There is a corresponding well-ordering in the motor centres. Each motor ganglion consists of the juxtaposition of motor centres, of which only the one belonging to the segment is the original; the others represent copies of the motor centres of the subordinate segments. Neighbouring segments project into neighbouring copies.
Further sorting took place when different types of receptors formed. We can assume that a new type of receptor usually formed from an existing type of receptor. Then the associated modality split into two new modalities. We assume that the projection neurons then also separated spatially.
The increase in receptor types was also associated with an increase in the number of receptors. Receptors of one type, such as tactile receptors, were distributed all over the body surface from a certain stage of evolution onwards.
The increasing number of receptors and types of receptors led to the transition from a knitted ladder-like nervous system to a tubular one. The reason was the tubular, worm-like body structure of these early animals. Thus, the body topology could be easily reproduced in the neural tube. The axons of the sensory projection neurons together with the axons of the motor projection neurons formed a neural tube, the interior of which was filled by a fluid-filled ventricular space. The half known as the posterior horn consisted of class 1, 2, 3, 4 and 6 neurons and their axons. The axons of the class 4 sensory projection neurons are surrounded by myelin in the human neural tube. This increases the range of the neuronal signals. The axons always run on the outside in the neural tube, where the white matter is located. The white colour is caused by the myelin.
Inside the posterior horn, on the one hand, there are the nuclei of the projection neurons of classes 1, 2, 3 and 4 and the unmyelinated axons of the neurons of class 3, which project to the motor neurons.
In the anterior horn of the neural tube are the nuclei of the motor projection neurons of classes 1, 2, 5 and 6 and their axons, which project to the motor neurons in the various segments. Between the anterior horn and posterior horn run the transverse connections formed by the class 3 and 2 neurons.
In addition to the anterior horn and posterior horn, there is also the lateral horn in the upper area, which lies between the two. It is formed by the cell bodies and axons of the autonomic neurons and belongs to the sympathetic nervous system.
A topological well-ordering is also observed in the neural tube. There is an order according to segments and according to modalities.
The projection axons of the class 4 neurons of the lower segments run internally. Higher, head-side segments attach their axons laterally on the outside.
In the posterior horn it is analogous. Axons of class 5 neurons, which project to tail-side segments, are located on the inside; here, too, higher, head-side segments attach their axons on the outside.
This results in a kind of tree-ring structure with regard to the afferent and efferent projections. Axons of the lower segments form the inner rings, while axons of the higher segments form outer rings.
But there is also a topological well-ordering with regard to the signal-providing receptors and the signal-receiving motor neurons. The receptors that supply signals to the sensory part, i.e. the posterior horn, developed gradually one after the other in the course of evolution. The axons that transport signals from the evolutionarily oldest receptors headwards are arranged on the inside, while those from the evolutionarily younger receptors are arranged on the outside. Thus, the different modalities form ring-shaped structures, which we call modality rings. A modality ring consists of a sensory half-ring, formed by the afferent projecting axons of class 4 neurons, and a motor half-ring. The neurons that form it receive their signals through the class 3 cross-connections, which receive input from the class 4 neurons.
In addition, the modal rings are divided into segmental rings. Inner segmental rings belong to deeper, tail-side segments.
We want to choose this basic structure of the neural tube as a starting model to analyse the future development of the central nervous system of vertebrates, mammals, primates and ultimately Homo sapiens.
Certainly, there were bilateria that never reached this neuronal structure. So it was possible that in many species the neuronal range of the signals could only overcome a few segments. Then the uppermost segment did not receive the complete information of the entire body from all segments, but only the input from one or two neighbouring segments. These animals could not become vertebrates in which the top segment had the complete input from all segments.
But the animals whose neural tube was constructed as described were still very dissimilar to vertebrates. In particular, all segments were still fairly identically constructed. Almost all organs were still present in almost all segments. In order to ultimately develop into a vertebrate, not only a spinal column had to develop. Many organs that were previously present in each segment had to regress so that they ultimately remained in only one or at least a few segments. We will call this process reduction. What could have favoured this development?
In my view, we owe this development to lateral inhibition. In each segment and in each of its neuronal nuclei, there were inhibitory interneurons that served to enhance contrast. This was achieved by the mutual inhibition of the neurons involved by means of the inhibitory interneurons. Inhibitory interneurons enabled neuronal competition. And with competition, there are winners and losers.
Let's look at the receptors of the sense of smell, also called the olfactory sense, as an example. It is one of the oldest senses. The multicellular organisms that later formed the segmented bilateria through colony formation already possessed olfactory receptors. No wonder that these were present in (almost) all segments in the early segmented animals, because in the beginning the segments were quite identical. Each olfactory receptor delivered its signal to a connected class 3 neuron as well as to a class 4 neuron. The class 3 neuron projected into a class 5 motor neuron, which contacted a motor neuron in that segment. In contrast, the class 4 neuron projected headward. In the neighbouring head segment, it again contacted a class 4 neuron, which projected to the next neighbouring head segment. Thus, this signal finally arrived in the head segment. There was already a vast number of sensory neurons there, which received the signals of exactly this modality from the most diverse segments. This is because each segment sent the olfactory receptor signals to all segments above it, including the head segment.
The neurons that received the signals in turn activated inhibitory interneurons that caused lateral inhibition. Thus, each olfactory signal inhibited the olfactory signals of the neighbouring neurons. All segments of the body were therefore in neuronal competition with each other with regard to olfaction. The strongest competition was in the head segment. This is where all the olfactory signals from all the segments arrived.
The signals from the tail segment were most affected by the lateral inhibition, because the olfactory signals from all the other segments had an inhibitory effect on them. This was also due to the fact that the olfactory receptors in the different segments were excited by a scent at the same time. At that time, the animals lived in the uro-ocean; a scent spread quickly in the water to all sides. Thus, all olfactory receptors in all segments that reacted to an existing scent were active at the same time. This was especially true when the animals' bodies were still relatively small.
Over a long evolutionary period, this led to neuronal atrophy. Neurons that are constantly suppressed regress and atrophy. At the end of the process, they are no longer present. Thus, the olfactory receptors in the tail segment disappeared completely. In the course of evolution, no olfactory receptors were created at all in the last segment, the genetic programme had changed.
However, the midline nuclei were also affected by this development. The olfactory midline centre, which I believe in humans is formed by the septal nuclei (nuclei septales), also no longer received input in the tail segment after the atrophy of the olfactory receptors, so it also atrophied.
This was not the end of the development, because now in the remaining segments - with the exception of the tail segment - all olfactory signals were again competing for dominance. Neuronal competition knows winners and losers. The winner was the head segment, the losers were the remaining segments. In the course of a longer evolutionary process, all olfactory receptors in all subsequent segments atrophied. Thus, the head segment had specialised in olfaction, the remaining segments had neither olfactory receptors nor did they have septal nuclei that functioned as olfactory mean nuclei.
But the neuronal competition was even more profound. Initially, the neurons were not yet as specified as in later evolutionary epochs. They did not yet differ significantly. Put simply, the neurons did not care at all which receptors they were excited by. At the beginning, there was no special specificity with regard to modality, or only to a small degree.
Therefore, in the head segment, all signals competed with each other, for example olfactory with visual, motor and others. Each signal competed with every signal in the same half of the body. But since the olfactory signals had already gained the upper hand (perhaps by chance - or because they were the first), the other modalities came under pressure. The lateral inhibition of the signals from each other and the greater assertiveness of the olfactory signals led to the receptors of the other modalities in the head segment being degenerated, atrophied and ultimately disappearing completely. Olfaction remained the winner in the head segment and remained as the sole modality in the head segment. The fact that olfaction in particular won the competition on the basis of lateral inhibition may also have been due to the fact that it was one of the earliest senses that the animal world had produced. The mean nuclei in the head segment could only receive olfactory signals. Mean nuclei that were supplied by other types of signals disappeared from the head segment.
It is not difficult to imagine that in the second segment the visual receptors won the multimodal competition of the segments and modalities. Thus, in the second segment, the visual sense became the sole winner, all other modalities disappeared here. And in all subsequent segments, the visual receptors would also have disappeared through atrophy, were it not for the vestibular sense. It is the reason why there was another pair of eyes besides the one in the second segment.
But those animals that had reached the stage of evolution described so far must have already possessed eye muscles. By default, there were six eye muscles in each eye.
Even spiders that belong to the simpler segmented animals have eyes. Here, a complete reduction of the number of eyes through lateral inhibition did not yet occur, so that several segments each have a pair of eyes. Most spiders have eight eyes. A distinction is made between main eyes and secondary eyes. The main eyes of the have eye muscles. While in vertebrates the eye muscles move the eye from the outside, in spiders the retina can be moved sideways with the help of the eye muscles. Many spiders have six eye muscles per main eye, but many species have fewer. This may already be a regression, because the sense of sight is of secondary importance in many spiders.
The possession of six (external) eye muscles in the segmented animals whose lineages led to the vertebrates is assumed here. There is a good reason for this.
In the third segment, which directly bordered the second, visual segment, the eye muscle signals were able to assert themselves and displaced all other receptor types of this segment, as well as the non-visual mean centres. Thus, in the third segment, a neuronal structure developed in the sensory centre, which we later call the tectum opticum, or tectum for short, in vertebrates. Here, visual signals overlap with the signals of the muscle tension receptors from the six (outer) eye muscles. The tectum developed into an important visual centre in vertebrates.
A seventh external eye muscle, the musculus retractor bulbi ("retractor of the eye") is missing in humans, but is developed in most other mammals. It is not used for eye movement per se, but to protect the eye when approaching obstacles.
The six outer eye muscles make it possible to always align the eyes in such a way that important visual objects can be well captured and analysed. The algorithm will be described later. The focal point module is used here as the principle module. Later, a difference module even makes motion vision possible. But more about that later. Here we first explain the step-by-step development of the segmented bilateria into vertebrates.
After the olfactory sense had established itself in the first segment, the visual sense in the second, and the receptors of the eye muscles in the third, the reduction continued through lateral inhibition.
Originally, there was a vestibular sense in each segment. It evaluated the position of the living being in space with the help of gravity. Even the simple multicellular organisms, which were not yet segmented, possessed it. Polyps, for example, used it to align their body axis vertically.
In a fluid-filled cavity, the statocyst, a grain of sand always rolled to the lowest point. The wall of the statocyst was lined with hair cells. These were bent by the weight of the grain of sand so that they generated action potentials. These activated the muscles that corrected the position of the body in the water so that the standard position was resumed. To do this, the tail of the body had to be bent accordingly. The muscles were activated by the action potentials of the statocyst.
This neuronal circuit necessarily required a signal crossing. The muscles on the contralateral side of the body always had to be activated.
When the segmented animals had evolved, each segment initially had two statocysts, one on the left and one on the right side of the body. The neuronal competition of all signals and all segments with each other led to the fact that in the first segment the olfactory signals, in the second the signals of the visual receptors and in the third the signals of the eye muscles had prevailed and displaced the other receptors in these three segments.
As a result, the statocysts in the first, second and third segments lost the competition and ultimately degenerated. From a certain evolutionary stage, there were no more statocysts in the first three segments. However, the statocysts in the remaining segments also entered the neuronal competition. Now the statocysts in the fourth segment most successfully inhibited the other statocysts, so that a gradual regression of the statocysts began in the remaining, tail-side segments. This took place over many hundreds, thousands or even millions of generations and was gradual.
Initially, the small opening through which a grain of sand first entered the statocyst became larger. The grain of sand fell out. The opening became even larger so that the water could enter the statocyst from the outside. Now, however, the hair cells no longer reacted to the weight of the grain of sand - this no longer existed. Instead, they reacted to the water flow caused by the movement of the animal. This was the beginning of the development of the lateral line system of aquatic animals. The water flow caused by the animal's own movements could be evaluated neuronally. Much more important, however, was the water flow caused by moving foreign objects. Predators caused such water currents when approaching, but also animals or plants that were on their own menu. Thus, the lateral line system became an important long-distance locating sense. It was especially useful in deep water that could not be penetrated by light. The lateral line system was present in all subsequent segments from the fifth segment onwards, while the vestibular statocysts only remained in the fourth segment.
Why did the neuronal competition of the lateral line sense receptors not lead to atrophy in all segments following the fifth segment?
Atrophy could only take place if neurons constantly receive similar (identical) signals. Only then are they always synchronously active at the same time and can also always inhibit each other at the same time. We call such modalities dependent modalities. Olfactory receptors, for example, which all deliver the same output signals, belong to the dependent modalities. However, the water flow in one segment was statistically different from the water flow in another segment, so that inhibition did not occur completely and at all times. Therefore, it did not lead to neuronal atrophy. There were sufficiently long periods of time when the signals from a lateral line receptor in one segment were not inhibited by other lateral line signals from other segments. We refer to such modalities as independent modalities. Almost all trunk modalities are independent modalities, because the dependent modalities only persisted in the head segments, and only in a single segment at a time. At least, that is the standard case.
The sensory and motor nuclei of the fourth segment developed into the torus semicircularis of vertebrates. In them, the signals of the vestibular sense were aligned with the trunk signals and thus ensured the desired orientation of the body in the earth's gravitational field.
We assign the signal crossing of the vestibular signals already described to the fourth segment. Here, the vestibular signals changed the side of the body. Since this was probably controlled by a change of the body side marker, all descending and ascending axons in the fourth segment switched equally to the contralateral side.
Thus, the descending visual signals in the fourth segment also switched to the opposite side. However, the eyes that followed the fourth segment before their later regression in every other segment did not show such a signal crossing. Therefore, their signals did not match those of the second segment. The lateral inhibition of the signals from the first pair of eyes could not work here. Therefore, the higher segmented animals, whose lineages lead to the vertebrates, have a second pair of eyes in the sixth segment. Its signalling pathways are uncrossed. However, the eyes of the subsequent segments fell victim to neuronal competition from lateral inhibition and were regressed.
Thus the vestibular sense was located on the fourth floor, the fifth was assigned to the lateral line sense. Here, too, it proved more favourable if the lateral line sensors excited the muscles of the opposite side, so that the body part concerned could be moved away from the point of impact. Therefore, here too, signal crossing became the standard and combined with the signal crossing of the fourth segment to form a general signal crossing for all ascending and descending signals.
The sixth floor contained another pair of eyes whose signal path was uncrossed.
We can certainly assume here, without contradiction, that in the early days of evolution it must generally have been compound eyes. Lenticular eyes developed from them in the course of evolution. But how could this process have been started in the future vertebrates?
A compound eye (with a sufficient number of facets) provides an upright and laterally correct image of the visual space on the associated receptor surface, which we can certainly also call the preliminary stage of a retina. There is no lens that turns the image upside down and displays it laterally reversed.
Let us now imagine that an edible object, a prey, is on the left side of the visual field. It will create an image in the compound eye on the left side of the retina, which activates the visual receptors. These now generate action potentials.
Let us now compare the signal path of the second segment with that of the sixth segment. According to our theory, there were two eyes in each of these segments. We look at the signal path of the left eye, because the prey can be seen on the left. In the beginning, the eyes sat sideways, there may not have been a common field of vision at all.
The visual prey signals from the left eye in the second segment activated class 3 neurons located in the sensory centre. These projected into the motor centre to class 5 neurons, which in turn projected tailward to the motor neurons of the trunk muscles. However, they had to pass through the signal junction in the fifth segment, so that they were now on the right side of the body. As a result, the visual signals caused the right trunk muscles to contract, and the animal's body bent and curved to the right. The resistance of the rear torso in the water caused the animal's head to move to the right as well. Thus, the head moved away from the prey. This was inexpedient.
Fortunately, there was still the pair of eyes of the sixth segment. Its signals did not have to go through a signal crossing. Therefore, they triggered a motor response in which the trunk curved exactly to the other side where the food was.
Thus, the visual signals from the second segment attempted a torso curve to the right, but those from the sixth segment attempted a torso curve to the left. They competed with each other.
If the eye signals from the second segment won, the prey swam away. If the eye signals from the sixth segment won, the prey could be eaten.
Those animals in which the eye signals from the sixth segment won the neuronal competition more often had a clear advantage. Thus, the eye signals of the sixth segment began to suppress the eye signals of the second segment. To do this, they moved headward on the axons of the sensory projection neurons of class 4 until they reached the sensory centre of the second segment. There they contacted inhibitory interneurons, which now began to inhibit the visual signals of the second segment.
This started the process of successive atrophy of the anterior pair of eyes. It lasted for many generations. The outwardly curved surface of the compound eyes in the second segment began to collapse and sink into the depths because the structures there also atrophied. It is even possible that a cavity developed where the ocular anlagen had previously been, because the atrophy was already showing consequences. It is not difficult to imagine that something like a primitive pit eye developed, because the visual receptors did not regress overnight, because the process of atrophy of an organ system took many generations.
However, the physical laws of light propagation caused pit eyes to produce a side-inverted, upside-down image. When these visual signals passed through the signal intersection, this twist was virtually cancelled out. The first image reflection around the image centre produced the pit eye, the second reflection caused the signal crossing. In this way, the two could neutralise each other. Now the visual signals of the second segment were usable again.
When the pit eye evolved by forming an outer, transparent protective layer in which a converging lens also formed, the imaging quality of the eyes in the second segment was significantly better than the imaging quality of those in the sixth segment.
The increase in the number of visual receptors in the new lens eyes of the second segment led to them winning the neuronal competition with the compound eyes of the sixth segment. Now one could have expected a complete atrophy of the eyes in the sixth segment.
However, this did not occur. The relatively simple eyes in the sixth segment had functioned as signal suppliers of the optical mean nucleus in the sixth segment during this very long evolutionary process. Here, the mean brightness was evaluated and controlled the day and night activity of the animal. This control mechanism was so important that it had to be maintained. Therefore, these rudimentary eyes took over the task of circadian control of the optical mean nucleus. In humans, we find this optical control centre, which controls activity during the course of the day, in the nucleus suprachiasmaticus.
The seventh segment was - according to this theory - the input and output segment of the primordial brain of these animals. Here, the signals of all receptors from all trunk segments arrived and were passed on to the head segments to interact with them. The output was in turn passed to the trunk segments.
The forebrain is thus formed by segments one to seven, which we call the head segments. The remaining segments are the trunk segments. It is possible that future research will expand this simple model by adding more segments.
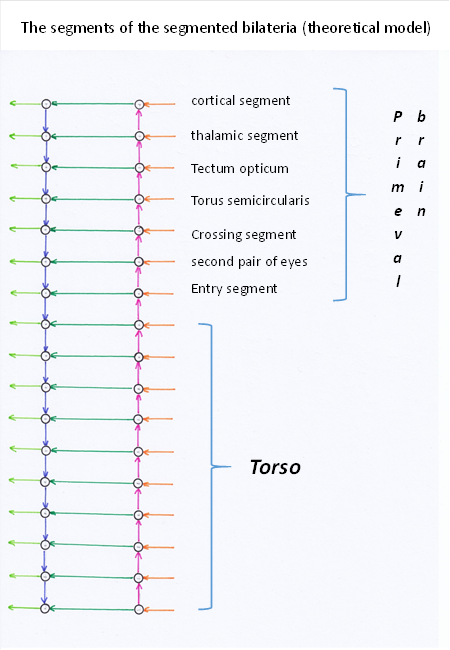
Figure 3: Theoretical model of the nervous system
The trunk segments - with the exception of the last trunk segment, the budding segment - were initially completely identical in structure. They all had the same internal (and external?) organs. But here, too, development continued.
Organs were neuronally controlled by the mean centres already described. However, the midline centres also participated in the cross-segmental signal exchange. Each midline centre also projected headward into all higher-level neighbouring segments via class 4 neurons and tailward into all lower neighbouring segments via class 5 neurons.
There was also neuronal competition between these mean signals, each arriving in the sensory and motor nuclei of each segment. Inhibitory interneurons provided the lateral neighbour inhibition.
In the course of a long evolutionary period, this resulted in a few of the many similar mean nuclei present in each segment becoming dominant, while atrophying in other segments.
The atrophy of a neuronal mean nucleus in a segment had the consequence that the organ or organ system it supplied in this segment also atrophied, i.e. gradually degenerated, since the control signals were absent. This process certainly took many millions of generations, especially since the result was ultimately manifested hereditarily.
In the end, there was no longer every organ in every segment, but certain organs only once on each side of the body. However, if this was not sufficient to sustain life, this organ could be preserved in several directly adjacent segments, but in the remaining segments its complete regression took place.
However, the animals at this stage of development were not yet vertebrates. They lacked the spinal column. But also in the neuronal field, something very important was still missing: the cerebellum.
In a monograph on the brain and consciousness, the origin of the spine need not necessarily be discussed. There is indeed the hypothesis that the chordates evolved from the tunicates, whose larvae already had a chorda and a neural tube, but let this field be left to others.
But how did a cerebellum develop? All chordates have a cerebellum, so they could well be called cerebellum animals.
In my monograph "Brain Theory of Vertebrates" I presented a theory of the origin of the cerebellum, which is reproduced here in short form.
The starting point is the bilaterality of the creatures considered here, which are regarded as the precursor of vertebrates. In these animals, the left and right halves of the body were in neuronal competition. Each neuronal centre of each segment projected via class 2 neurons into a side-changing nucleus. There, the signals were switched to an excitatory projection neuron that responded to the body marker of that side of the body. Its axon moved directly to the similar neuronal centre, which, however, was located on the opposite side. There it activated inhibitory interneurons, which caused a lateral inhibition of the neurons there. Thus, both halves of the body were in neuronal competition with each other.
Stronger signals on one side of the body prevailed over weaker signals on the other side. Depending on whether food was smelled, seen or felt on the right or left side, the right side of the body was able to approach the food and grab it so that it could be eaten. It was also possible to avoid predators, even if we do not always understand the algorithms for this yet.
The cerebellum developed in the seventh segment. This was also the input segment of the primordial brain of our animals. The sensory centre received all sensory signals from all trunk segments. This input nucleus split into two sub-nuclei in vertebratescalled the nucleus cuneatus and the nucleus gracilis. The first receives the receptor signals from the upper half of the trunk, the other from the lower half. At the same time, this segment is the output floor on the motor side.
The motor centre of the seventh segment became the nucleus ruber, whose output drives the motor neurons of the trunk and is thus responsible for the animal's movements.
The contralateral inhibition of the opposite side, which occurred in each segment in the initial period, was realised by the axons of the class 2 neurons moving to the associated side-switching nucleus. There they were switched to a side-switch neuron, moved to the opposite side and ended in the corresponding neuronal nucleus on inhibitory interneurons. The motor side-switch nucleus of the seventh segment is called the nucleus olivaris. It receives the output of the nucleus ruber and projects excitatory into inhibitory interneurons of the contralateral nucleus ruber in early primitive time. Thus, contralateral inhibition is realised at the brainstem level in early evolution. We count segments 2 to 7 as part of the brain stem, segment 1 is the cortex.
The inhibitory interneurons of the nucleus ruber, which were used for this contralateral inhibition, spatially separated from the nucleus ruber during evolution and formed their own nucleus. When neurons specialise, they come together to form their own nuclei (or layers). This new nucleus of inhibitory neurons took the excitatory output of the contralateral nucleus olivaris and switched it to the inhibitory transmitter GABA. The axons of its neurons moved to the nucleus ruber and inhibited their associated neurons there in a point-to-point mapping. Thus, each neuron of the nucleus ruber inhibited the mirror-image neuron of the nucleus ruber of the opposite side.
This new nucleus, consisting of GABAergic neurons, is what I call the nucleus Purkinje in my monographs. Its neurons are the original form of Purkinje cells. In the early developmental phase of the cerebellum, they served the contralateral inhibition of the nucleus ruber of the opposite side, their input came from the olive nucleus.
When a type of neuron in a neuronal nucleus specialises in a specific, new task, these neurons often separate from the previous nucleus and form their own, new nucleus. This can be regarded as a descendant of the old nucleus. In this respect, the nucleus Purkinje is a descendant of the inhibitory interneurons of the nucleus ruber.
This was the first step towards a cerebellum. All that was missing was the modification of the output. As is well known, the output of Purkinje cells ends in a neuron nucleus consisting of excitatory neurons. Now, neurons with the neurotransmitter glutamate, if they receive inhibitory input, can only deliver an excitatory output if they are additionally excited by external signals. The supplied excitation must be stronger than the supplied inhibition for this to happen. Which nucleus could provide the necessary excitation here?
In the seventh segment, only the glutamatergic mean nucleus can be considered. This nucleus was the formatio reticularis (or its glutamatergic part). It received (at that time) the mean value input from the sensory and motor nucleus, i.e. from the nucleus ruber, the nucleus cuneatus and the nucleus gracilis. All the signals were related to motor function. The seventh segment specialised in this.
Averaging kernels integrate as many available signals as possible. Therefore, the signals from the Purkinje cells were integrated into the formatio reticularis. Here, a new type of signal processing occurred. The neurons of the nucleus reticularis were permanently excited (during movements) because they represented the mean value of the motor excitation. If a Purkinje cell inhibited this average excitation, a residual signal remained. If the inhibition signal was strong, the residual signal was weak. If, however, the inhibition signal of the Purkinje cell was weak, the residual signal in the mean value neuron was strong.
This corresponded to the inversion of the signal strength, we will call this inversion. A strong signal is transformed into a weak one by inversion, but a weak signal into a strong one. The monotonicity, i.e. the signal strength curve, is reversed.
This inverted output was excitatory. Its motor target was still the nucleus ruber. Each motor signal from one half of the body was now converted into an inverted signal and excited the motor counterpart on the opposite side. This is called co-activation in the neurological literature.
Those mean neurons that received the output of the Purkinje cells separated from the nucleus reticularis and formed an independent nucleus of their own. This consisted of glutamatergic neurons that received mean excitation from the nucleus reticularis. At the same time, they were inhibited by the Purkinje cells. The excitatory output of this nucleus led to the inverse excitation of the contralateral nucleus ruber.
This nucleus became the first cerebellar nucleus of the original cerebellum. The Purkinje nucleus now represented the cerebellar cortex, and in its inner medullary canal was the cerebellar nucleus, whose output supplied the nucleus ruber with inverted signals.
This early cerebellum inverted the motor signals and fed them to the motor counterparts. Previously, a muscle was tensed, but its counterpart was inhibited contralaterally. Now - with cerebellum - co-activation of the counterparts was achieved. The counterpart muscle was no longer completely slack, but also had a certain basic tension. This brought firmness into the body. This made it possible for the first animals to leave the water habitat. The co-activating muscles compensated for the gravitational stress that occurred. Without co-activation, movement on four legs on land would have been impossible. In this respect, the spinocerebellum is the precursor of the land gait of aquatic animals. (The development of the vestibulocerebellum has been described in detail in my previous monograph and can be read there).
This original cerebellum already received three types of input signals from the beginning, but initially exclusively via the climbing fibre projection from the olive of the opposite side:
1. Input signals from the vestibular nucleus of the fourth segment (neuron class 5)
2. Motor input from the seventh floor of the nucleus ruber (neuron class 5)
3. Mean input of the seventh floor from the nucleus ruber (neuron class 6)
This led to the tripartition of the cerebellum into a vestibulocerebellum, a spinocerebellum and a pontocerebellum, likewise the cerebellar nucleus divided into three subnuclei, one of which was even divided into two.
While neuron class 5 provided the input for the vestibulocerebellum and the spinocerebellum, the mean neurons of class 6 from the nucleus ruber supplied the pontocerebellum. This was relatively insignificant at the time because the number of mean signals was relatively small. It served the inverse activation (co-activation) of the mean nuclei of the opposite side.
Only with the development of a mossy fibre projection into the cerebellum could the pontocerebellum become a learning machine, as John C. Eccles already suspected. This will be explained later, but can also be read in the previous monograph.
The further development of the cerebellum began with the formation of a mossy fibre projection. This moss fibre projection was possible because the cell bodies of the neurons in the neural tube were always arranged on the inside, while on the outside the axons ran head and tail.
In the cerebellum, the Purkinje cells received the signals from the opposite side via the climbing fibres of the olive, while the signals from their own side travelled upwards in the axons of the headward-projecting class 4 neurons. Similarly, the sensory neurons of class 3 and 4 were in the immediate spatial vicinity of the Purkinje neurons.
Already in the neural tube and also in the spinal cord, the tactile and pain signals acted on the motor neurons and inhibited them. This resulted in self-protection of the animal. This inhibitory effect was transferred to the cerebellum. It was realised there by interneurons, which we call star cells and basket cells. The signals from the tactile and pain receptors reached the granule cells via the moss fibres, whose axons excited the stellate and basket cells. These in turn inhibited the Purkinje cells. These could no longer inhibit the output neurons in the cerebellar nucleus as strongly, so that they fired more strongly. The excitatory signals for this came from the associated median nucleus, the formatio reticularis. The output neurons of the cerebellar nucleus in turn excited the contralateral motor neurons so that the muscles contracted and moved the affected body part away from the site of pain.
When very strong tactile or pain signals from the ipsilateral side reached the cerebellum, they hit the granule cells of the cerebellum via the mossy fibre projection (class 3) and inhibited the Purkinje cells via the star and basket cells. The more strongly inhibited Purkinje cells now inhibited the output neurons in the cerebellum nucleus significantly less, so that the latter now provided a stronger output signal due to their mean value excitation supplied by the formatio reticularis. As this output reached the contralateral muscles, they contracted and moved the body part away from the site of pain. For example, the arm was pulled away when a hot cooker top heated it. In this way, the spinocerebellum took over self-protection. This had previously existed at the spinal cord level; it is called the spinal cord's own apparatus. In the course of the development of the mossy fibre projection, the spinocerebellum took over the protective functions of the body and the spinal cord's own apparatus, which was now only exercised there by the spinal cord in emergency situations, since the cerebellar pathway was time-delayed.
Later, when the number of signals from the animals increased explosively because a myriad of receptors were formed, the size of the cerebellum increased enormously. It could no longer remain inside the neural tube. This was also because the number of feeding axons of the climbing fibres, which always had to pull on the outside of the neural tube to the cerebellum, increased enormously.
The severe space-occupying lesion caused the neural tube to rupture at the level of the cerebellum. Through the opening created, the cerebellum penetrated to the outside. The ventricular fluid that filled the inside of the hollow neural tube was also able to spread outwards. Since then, the brain has also been washed around from the outside by the ventricular fluid. This now also acts as a shock absorber.
This reversed the order of the neuron layers. The neurons of class 1 are located in the neural tube on the inside of the ventricular wall, then neuron classes 2 to 6 follow in this order from the inside to the outside. In the cerebellum and cortex, the neurons of class 1 also seek the vicinity of the ventricular wall, but there the brain is surrounded from the outside by the ventricular fluid. Therefore, the neurons of class 1 in the cortex are on the outside, those of class 6 on the inside. Thus the cortex has the following neuron stratification from the outside to the inside:
1. Class 1 neurons, top, outside
2. Class 2 neurons
3. Class 3 neurons
4. Class 4 neurons
5. Class 5 neurons
6. Class 6 neurons, bottom, inside
We should bear in mind that actually all neuron nuclei have such stratification, even if the stratification sequence in neural tube was exactly the opposite. But the head floors (at the end of a remodelling process) certainly had the same stratification as the cortex. Especially the thalamus, which was located in the second segment of the original cord ladder nervous system.
The neurons of class 6 are mean value neurons. The human brain and also the vertebrate brain owe to them the ability for abstract signal analysis and the storage of complex signals. This assertion is proven in this monograph, and there is also corresponding proof in my earlier monographs.
Without mean neurons we would - as will be proven later - not be able to recognise the shape of objects, store them in our brain and recognise them later. We would only have a very limited sense of the body. We know instinctively what angles the various joints of our body form, or what position we occupy in the earth's gravitational field. Likewise, we are able to estimate the movement and speed of objects of all kinds. We owe all this to the existence of mean neurons. These magnocellular neurons are found in many substructures of the brain, for example as Purkinje cells in the cerebellum, as pyramidal cells in the fifth and sixth cortex layers, as GABAergic striatal neurons in the basal ganglia system, but also in the magnocellular part of the nucleus ruber or in the nucleus subthalamicus. The functioning of the brain modules that use such magnocellular mean neurons will be analysed from an information-theoretical perspective in the next chapter.
Just as significant as the median neurons are the median nuclei of the brain. Originally present in every segment, they serve to control important vital functions. Due to the neuronal competition between the signals of all segments among each other, a reduction through atrophy also occurred here, so that important mean value nuclei only remained in a few segments, often only in a head segment. In the course of evolution, the median nuclei took on important, completely new tasks. We owe our short-term memory and the ability to analyse movements and directions of movement to the dopaminergic median nuclei. We owe the glutamatergic median nuclei, among other things, the ability to learn complex signals. All this will be demonstrated in this monograph. We list the most important mean nuclei of the human brain below, but we should bear in mind that they were all already present in the vertebrate brain.
Mean nuclei of the brain:
1. reticular formation
2. Substantia nigra pars compacta and area tegmentalis ventralis
3. Subthalamic nucleus
4. Septal nuclei
5. Raphe cores
6. Locus coeruleus
7. Magnocellular part of the basal nucleus of the amygdala
The layered concept of the brain was constantly developed along the evolutionary path from lower vertebrates to mammals and especially to Homo sapiens. The further development was also a consequence of the further development of the receptors. In the course of evolution, new types of receptors were constantly developed. Some of them were new developments, for example the muscle spindles, others emerged from already existing receptors, for example through gene duplication. These new types of receptors enabled the associated modality to analyse the environment and also its own body ever more specifically, making the environmental and body images much more meaningful. The new types of receptors led to the splitting of the modalities.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan